
פרויקט ומחקר חדש ברפואת רשתות ואפיגנטיקה.. הפעם אני מתמקד באריכות ימים של רקמת החיבור והמטריצה החוץ-תאית (The Extracellular Matrix – ECM), המוגדרת כאחד ה-Hallmarks of Aging. המחקר מסייע ובתחומים קליניים מתקדמים (ריפוי פצעים, פיברוזיס ריאתי/כבדי, קרטילג', מטריצות דה-צלולריות בהנדסת רקמות) קשובים פמחקר החדש – כבר משתמשים בגישות מבוססות-ECM או מכוונות שיקום המטריצה החוץ תאית–ECM [מקור2, מקור3].
הגענו להישגים פנומנליים בקליניקה בשיקום סוכרת סוג 2 (תוך 3 ימים עד חודש), שזהו הבסיס ההכרחי לפני שיקום כליות או לב. על ההישגים בהפעלת רשת החלמה בקרב חולי כליה פרסמתי מאמרים רבים, שנקשרים יותר ברפואת העתיד ובתובנות שהמחקר מקדם. כעת, אני רוצים לפצח לעומק את ה-ECM, נושא שטרם חרשתי במלואו מעבר להפעלה בסיסית של ה-NRF2 באמצעות נבטי ברוקולי וסולפוראפן – כאן
אציג לפניך את:
עקרונות היסוד המדעיים והמטאפוריים שגיבשנו כבסיס
ואסיים עם
הפתרון הרשתי המורחב: כדי לשקם את ה-ECM, הפעלת NRF2 לבדה (בלימת חמצון) אינה מספיקה. יש לשלב אותה עם הפעלת אוטופגיה (Autophagy) לפינוי ה-AGEs מחלבוני הרקמה, ואספקת חומרי הזנה ספציפיים (כמו פוליפנולים מזרעי ענבים 🍇 וליקופן 🍅) לבנייה נקייה [מקור].
קריאה מהנה
המדע הגדיר רשמית את ה-ECM כאחד ה-Hallmarks of Aging (סימני ההיכר הרשמיים של ההזדקנות). זהו כבר לא נושא שולי ברפואה או "רקמת תמיכה" פסיבית, אלא המפתח לאריכות ימים ברמת הרשת הגופנית ונושא מוביל במרחבה של רפואת העתיד [מקור3].
- שומרי האוצר הזעיר – רפואת העתיד – חלק א' / ירון מרגולין
- שומרי האוצר הזעיר – חלק ב' רפואת העתיד / ירון מרגולין
- מסלול איתות Wnt

המטריצה החוץ-תאית (ECM) היא האוקיינוס הפנימי שבו חיים כל תאי הגוף. כשהיא גמישה ונקייה – התאים מתפקדים היטב. כשהיא מתקשחת ומתלכלכת (בעיקר בגלל AGEs מסוכר עודף, דלקת וקורטיזול גבוה) – התאים נכנסים למצב דלקתי כרוני והזדקנות מואצת.
הפתרון אינו רק "לתקן את התא", אלא לנקות ולשקם את הסביבה שלו: הפעלת אוטופגיה, עיכוב יצירת AGEs (קרנוזין + חומצה אלפא-ליפואית), שיפור ניקוז לימפטי, הורדת קורטיזול (אדפטוגנים) והפעלת NRF2 יחד עם חומרי בנייה.
שיקום המטריצה הוא מפתח מרכזי לגמישות, תפקוד ואריכות ימים. ירון מרגולין
האוקינוס הקדום והנוזל הבן תאי – המתריצה
לפני מיליארדי שנים נולדו החיים במים.
מאז השתנו היבשות, נכחדו אינספור מינים, הופיעו יערות, דינוזאורים, יונקים ואדם.
אבל יש דבר אחד שכמעט לא השתנה. כל תא בגופנו עדיין חי בתוך ים, שוחה כדג.
אוקיינוס פנימי, זעיר, שקוף ושקט – המרחב שמקיף את התאים שלנו. המדע קורא לו המטריצה החוץ־תאית (Extracellular Matrix, ECM).
החוקרים מעלים אפשרות חדשה לגבי מרחב הקיום הזה, האם הסיפור החשוב אינו התא עצמו אלא האוקינוס שבו הוא חי.
מתוך שאלה זו נובעת השאלה שמענינת את אריכות הימים, את ביוהאקינג העוסק בהארכת חיים בחברה העשירה ביותר בעולם ואת מבקשי ההחלמה, והפעלתם של נתיבי ההחלמה הביולוגיים שפועלים בגוף האדם. שאלת האוקינוס הפנימי (רקמת החיבור והמטריצה החוץ-תאית (The Extracellular Matrix – ECM)) קשורה בירידה בתפקודם של חלבוני ההחלמה, ומחסור בשומרי האוצר הזעיר – נושא מרכזי ברפואת העתיד, מה שמכונה בהמשגה כוללת, הגד שמרכז הרבה העדר תפקוד תאי, הזדקנות. האם הזדקנות אינה מתחילה רק בתוך התא. ייתכן שהיא מתחילה כאשר האוקיינוס שסביבו משתנה – מתקשה, מתלכלך, מאבד את גמישותו ומפסיק לדבר בשפת החיים הבריאים ❓
נצא הפעם עם ירון מרגולין למסע אל אותו ים נשכח. מסע אל הרקמה שבין התאים. אולי, גם מסע אל אחת ההתחלות העתיקות ביותר שעדיין חיות בתוכנו.

לפעמים אני חושב על המטריצה החוץ־תאית כמו על מפרץ שקט.
כל עוד המים צלולים, בצבע תורכיז כל יצור שחי בהם מקבל את מה שהוא צריך: חמצן, מזון, מסרים וסביבה בטוחה.
אבל די בכך שמי ביוב יזרמו אל המפרץ כדי שהכול ישתנה [מקור].
המים עדיין שם, היצורים עדיין שם, אך הם חיים בעולם שזוהם בחוסר אחריות מוכר למי שמכיר את התנהלותם של שרי איכות הסביבבה [צחי הנגבי, ויהודית נאות (הזרמות הבוצה של ה"שפד"ן" – במשך שנים ארוכות, מפעל השפד"ן הזרים מיליוני קוב של בוצה (ביוב מעובד) ישירות ללב הים התיכון מול חופי ראשון לציון ופלמחים), גילה גמליאל (אסון הזפת), אבי גבאי, גלעד ארדן (דליפת הדלק בנחל צין ובמפרץ אילת), תמר זנדברג ועידית סילמן (זיהומים חוזרים בנחלי הצפון). הרעלים מצטברים, התקשורת משתבשת, והחיים הופכים למאבק הישרדות.
ייתכן שגם הרקמה שלנו פועלת כך.
כאשר תוצרי גליקציה (AGEs), תוצרי פירוק של המטריצה, עודף רדיקלים חופשיים ואותות דלקת מצטברים במרחב שבין התאים, התא נותר שם, אין לו דירת מפלט, ואין לו לאן לברוח, רק מפני ששר איכות הסביבה בז לה או משהו התקלקל במרחב חייו. הוא מגיב על ההתרחשות באמצעים העומדים בפניו, מגיב מול העולם שבו הוא חי שהשתנה בגלל הזנחה פושעת [מקור1, מקור2].
אולי אחת השאלות החשובות ביותר של רפואת העתיד אינה רק כיצד לתקן את התא, אלא כיצד לנקות את הים שבו הוא חי.
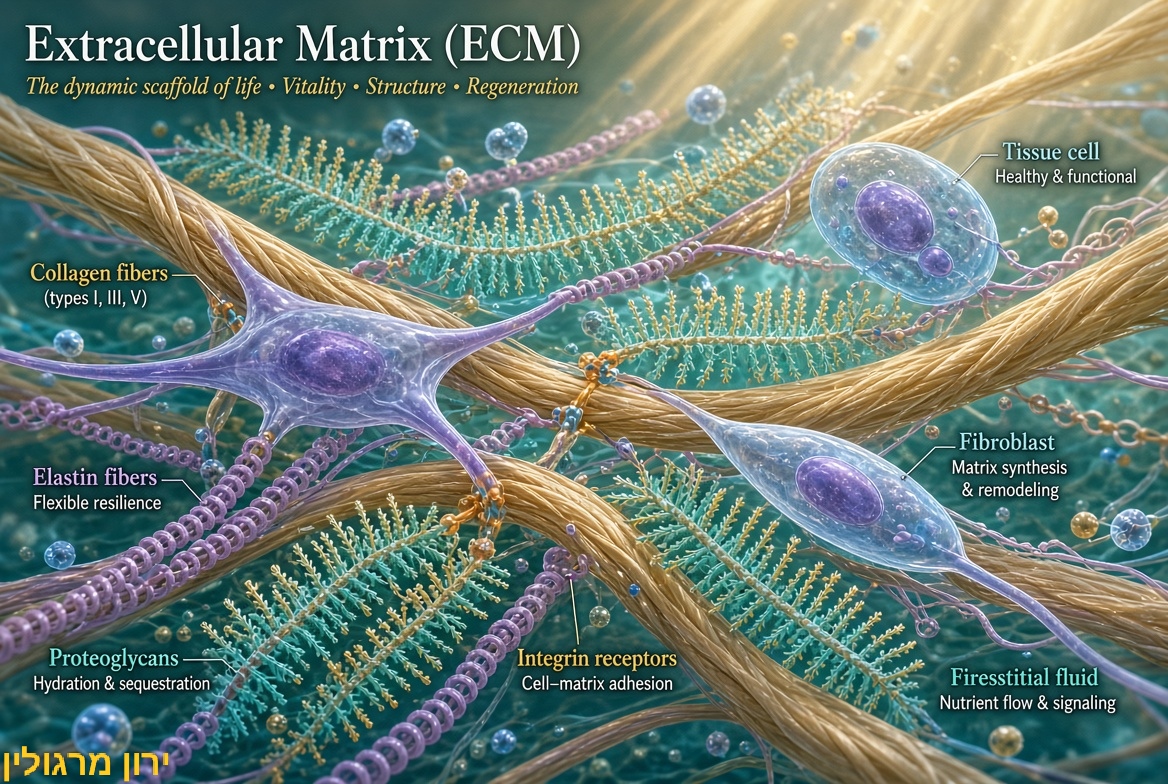
מהי המטריצה החוץ-תאית (ECM) הסימפוניה של הביולוגיה?
אם התאים הם הנגנים, ה-ECM הוא האולם שבו הם מנגנים. האולם הזה מורכב מרשת תלת-ממדית של פיברונקטין, קולגן, אלסטין ופרוטאוגליקנים.

כל רקדן יודע שהקסם מתחיל כשכל הקהל נושם איתו, כשהוא נע על הבמה. חוויה שבה השקט נחווה בכולם וגם התאוצה והדימינואנדו. אני שבתי וחוויתי זאת כשהופעתי ב"מחולות בני ברית דוד" באולמות בישראל ובבתי אופרה באירופה. זו החוויה שמעוררת את הקהל לעמוד על הרגליים בסוף המופע בקריאות עידוד והכרת תודה שמחייבות להעניק לו בחזרה לפעמים עד 5 הדרנים…
התגלית המדעית המטלטלת היא שהתאים מקשיבים לאולם. דרך קולטנים שנקראים אינטגרינים (Integrins), התא חש פיזית את הלחץ, המרקם והקשיחות של הסביבה שלו, ומתרגם את האות המכני הזה לאות כימי שמשנה את ביטוי ה-DNA בגרעין (תהליך שנקרא Mechanotransduction). [מקור1, מקור2, מקור3, מקור4, מקור5, מקור6].
כשמציבים תא שנאבק בגורמי הרס, וחלש בתוך מטריצה (ECM) של גוף צעיר, התא המכונה במחקר זקן עובר יובנליזציה (הצערה) וחוזר לתפקד כמו תא צעיר – המיטוכונדריות שלו משתקמות והוא מפסיק להפריש חומרים דלקתיים!
כלומר, הזדקנות האיברים היא מושג אופנתי, אבל קשור בתפיסתה של הרפואה הישנה, תלוית תרופות לתחזוקת המחלה, ומי שנהנה מחוסר סקרנות לקדם מסלולי החלמה. בפועל תפקוד התא (הרענן/לא זקן) מוכתב במידה רבה על ידי מצבו של האוקינוס הפנימי, מצב ה"אולם" ולא רק על ידי הבמה בה התא עצמו פועל, בעזרת נאמניו [מקור1, מקור2, מקור3, מקור4].
המטריצה החוץ-תאית (ECM) היא הרשת התלת-ממדית של חלבונים (קולגן, אלסטין, פיברונקטין), פרוטאוגליקנים ומולקולות נוספות שמקיפה את התאים. היא לא רק "פיגום" מכני אלא גם:
משדרת אותות מכניים וביוכימיים לתאים (mechanotransduction).
משפיעה על התמיינות, חלוקה, הישרדות והזדקנות של תאים.
משתנה עם הגיל: מתקשחת, מתפרקת, מצטברות בה קשרי צילוב (cross-links [מקור]) כמו AGEs, והרכב שלה משתנה [Nature, Aging and Disease, Cell –כאן].
שלושת אויבי הרשת של המטריצה
כדי לכתוב על "הכאוס המולקולרית מה קורה לאולם כשאנחנו מזדקנים" ?
עלינו להכיר את שלושת האויבים המרכזיים של ה-ECM, שהופכים אולם קונצרטים מפואר למחסן חלוד, וחוף ים מזוהם, מסוכן לרחצה:
- ההתקשחות וההצלבה (Advanced Glycation End-products – AGEs):
עודפי סוכר (שאנחנו מכירים היטב מתאבת המתוק, הפחמימות, יתר טריגליצרידים והסוכרת [מקור1, מקור2, מקור3, מקור4, מקור5]) נקשרים לקולגן ולאלסטין בתהליך של גליקציה. הם יוצרים קשרים צולבים (Cross-links) שהופכים את רקמת החיבור לקשיחה ולא גמישה. ההתקשחות הזו משדרת לתאים דרך האינטגרינים אות קבוע של: "אנחנו תחת מתקפה, תפעילו דלקת!" [מקור1, מקור2 , מקור3, מקור4, מקור5]. תאי הגוף (כמו פיברובלאסטים שמייצרים את רקמת החיבור) "מרגישים" את הסביבה שלהם [מקור]. במצב תקין, רקמת החיבור (המטריצה) היא גמישה, רכה וקפיצית. כאשר עודפי סוכר נקשרים לקולגן (תהליך הגליקציה שמייצר AGEs), הם הופכים את הרקמה לקשיחה, נוקשה ומתוחה, כמו גומי ישן שהתייבש בשמש. הגוף מתקשה, הגמישות אובדת. לתאים יש זרועות קטנות שנקראות אינטגרינים (Integrins). האינטגרינים נוגעים ברקמה הזו כל הזמן. כשהאינטגרינים מרגישים שהסביבה מאבדת מגמישותה, והופכת לקשיחה ומתוחה מדי, התא מפרש את השינוי הזה כלחץ הפיזי שמופעל עליו, לכאורה בגלל פציעה, קריעה או חדירה של גורם זר (מתקפה). 2. תרגום האות המכני לאות כימי. ברגע שהתא מרגיש את הקשיחות הזו, הוא לא יודע שמדובר "רק בסוכר", שהאדון דחף אליו בכוח התאווה למתוק. מבחינתו, הלחץ הפיזי אומר "יש כאן מצב חירום". האינטגרינים נזעקים מיד ושולחים אות חזק לתוך גרעין התא. האות הזה מפעיל את מתגי האפיגנטיקה הדלקתיים (כמו פקטור השעתוק NF-κB), והתא מתחיל להפריש חומרים דלקתיים (ציטוקינים) כדי "להילחם" באיום הפיקטיבי. בשורה התחתונה: הקשיחות הפיזית שנגרמת מהסוכר (AGEs) מטעה את התאים. היא גורמת להם לחשוב שהם נמצאים תחת מתקפה פיזית מתמדת, מה שמאלץ אותם לייצר דלקת כרונית (Metaflammation) שלא נפסקת אף פעם – פשוט כי הרקמה לא משתתפת בשיעורי הגמישות של הרקדן ונשארת קשיחה. בנוסף הלקוח והרופא שלו, לא שוקלים דבר פרט לתרופה, כשהשכל הישר זועק, תפחית סוכר ופחמימות! אבל שכל זה הוא חלק מרפואת העתיד. - אנזימי ההרס – ה-MMPs (Matrix Metalloproteinases):
אלו "פועלי ההריסה" של הגוף. במצב תקין הם מנקים קולגן ישן. במצב של דלקת כרונית (Metaflammation), הם יוצאים מאיזון ומפרקים את הרקמה לחתיכות [מקור1, מקור2, מקור3, מקור4]. - השברים הרעילים (ECM Degradation Products):
כאשר ה-MMPs מפרקים קולגן ואלסטין בצורה פרועה, נוצרים שברי חלבון קטנים (כמו אוליגו-פפטידים מאלסטין). מחקרים דרמטיים מראים שהשברים האלו: שברי אלסטין המשתחררים למחזור הדם בעקבות פירוק המטריצה החוץ-תאית (ECM) מפעילים באופן ישיר מונוציטים ומקרופאגים (תאי חיסון). ההפעלה הזו מתבצעת דרך קומפלקס קולטנים ספציפי בתא (הכולל את החלבון NEU1). תהליך זה גורם לתכנות מחדש של מערכת החיסון, מעורר דלקתיות מערכתית כרונית (Inflammaging) ומקצר באופן מובהק את תוחלת החיים. הפסולת של הרקמה הופכת לרעל [מקור1, מקור2 , מקור3 מקור4].
כדי "לסנן" ולטהר את האוקיינוס הזה, רפואת הרשתות מציעה פרוטוקול משולב שמורכב מכמה פעולות ממוקדות
לנקות את האקווריום
אם הדגים באקווריום חולים, רוב האנשים ינסו להזריק להם תרופות ישירות לגוף (רפואת התרופות הישנה והסימפטומטית). אבל הרופא העתידי יבין בהשפעת מחקר מתקדם שהבעיה היא לא בדגים – אלא במים. הסוכר מתפריט התזונה שלנו הוא שהפך את המים הצלולים של האקווריום לבוץ דביק וחומצי. שיקום הגוף דורש מאיתנו קודם כל לטהר את המים: להפעיל את הפילטרים הטבעיים (אוטופגיה), לפרק את האבנית (AGE-breakers), למנוע הזרמת אשפה אליהם, ולהזרים מים נקיים כדי שהדגים (התאים) יוכלו לנשום ולרפא את עצמם." [מקור1, מקור2, מקור3]
נעבור עתה אל הדרכים האפשריות לשימוש בבואנו לנקות את האקווריום – את הנוזל החוץ תאי, את האוקינוס בו שוחים תאי גופנו, כדי שיתפקדו כראוי גם לאחר פגעי הסוכר, והצטברות ה-AGEs (תוצרי הגליקציה), הוא הופך לצמיגי, דלקתי וחומצי, מה שחונק את התאים ומונע מהם לתפקד. [מקור1, מקור2, מקור3].
הערה: תוצרי גליקציה מתקדמים (AGEs) מצטברים במטריצה החוץ-תאית (ECM) עקב תגובה ספונטנית בין סוכרים חופשיים לחלבונים ארוכי-חיים כמו קולגן ואלסטין. הצטברות זו יוצרת קשרים צולבים (cross-links) חריגים, מעלה את נוקשות הרקמה, פוגעת בתפקוד התאי, ומאיצה תהליכי הזדקנות וסיבוכים כלי דם בסוכרת. [מקור1, מקור2]
תיקון המטריצה החוץ-תאית
רפואת הרשתות אשר פותחת את הדלת לרפואת העתיד – מציעה פרוטוקול חכם, שנובע תמדרכי חשיבת דור ההיטק דרך שמשלב כמה פעולות ממוקדות:
1. הפעלת ה"צוללות" של האוטופגיה החוץ-תאית
extracellular matrix remodeling או macrophage-mediated clearance of ECM components.
אנחנו רגילים לחשוב על אוטופגיה 🧹 כמנגנון ניקוי בתוך התא. אולם, מחקרי מפתח מראים שתאי רקמת החיבור (פיברובלאסטים ומקרופאגים) מסוגלים להפריש אנזימים מיוחדים אל הנוזל החוץ-תאי, "לבלוע" את חלבוני הקולגן המזוהמים בסוכר (AGEs), ולפרק אותם בתוך הליזוזומים שלהם. [מקור1, מקור2, מקור3, מקור4, מקור5, מקור6]
- איך מפעילים את זה? באמצעות צום סירוגין מבוקר (Intermittent Fasting) ופעילות גופנית אינטנסיבית קצרה (HIIT). מחסור זמני באנרגיה מאותת למנהל העבודה AMPK, שמפעיל את מתגי הפירוק והבליעה של חלבוני המטריצה הפגומים [מקור1, מקור2, מקור3, מקור4, מקור5, מקור6].
2. שימוש ב"מפרקי קשרים" טבעיים (AGE-Breakers)
הסוכר מייצר קשרים כימיים קשיחים (Cross-links) שמדביקים את סיבי האלסטין והקולגן זה לזה והופכים את הרקמה לנוקשה. כדי לפרק את המנעולים האלו בצורה נגישה: [מקור]
- חומצה אלפא-ליפואית (Alpha-Lipoic Acid): נוגד חמצון ייחודי הפועל הן בסביבה מימית והן בסביבה שומנית [מקור]. היא הוכחה כמסייעת להפחתת עקה חמצונית המאיצה יצירת AGEs, ומשפרת את המטבוליזם התאי. [מקור1, מקור2, מקור3, מקור4]
- קרנוזין (carnosine): די-פפטיד טבעי (המצוי ברקמות שריר או כתוסף) הפועל כ"מגן הקרבה" (או "קמיקזה"). ברפואת הרשתות, הקרנוזין אינו מתפקד כנוגד חמצון רגיל, אלא כ"מגן הקרבה" פיזיולוגי (Sacrificial Shield – הוא נקשר לסוכרים תגובתיים ומונע מהם לתקוף את הקולגן, אך לא שהוא "מפרק" קשרים צולבים שכבר נוצרו.). כאשר האוקיינוס החוץ-תאי מוצף במולקולות סוכר הרסניות המחפשות להחריב ולהקשיח את סיבי הקולגן והאלסטין (תהליך הגליקציה), הקרנוזין מציב את עצמו בחזית כפיתיון כימי [מקור]. המבנה המולקולרי הייחודי שלו מושך אליו את הסוכר בעוצמה רבה יותר, והוא מתנדב לעבור את תהליך הגליקציה [מקור1, מקור2, מקור3]. הקרנוזין בולם, על ידי הקרבת עצמו, את היווצרות ה-AGEs על גבי רקמות החיבור, מונע את התקשחות האיברים, ומאפשר למערכת הניקוז התאית לשטוף את "הסוכר השבוי" החוצה בבטחה [מקור]. מרגע שהקרנוזין מושך אליו את מולקולות הסוכר הפראיות שבנוזל החוץ-תאי, (נקשר אליהן במקום שהן ייקשרו לקולגן) הוא מונע את התקשחות האוקיינוס התאי. [מקור1, מקור2, מקור3].
- סינטזה של קרנוזין (carnosine), כמו סינטזה של גלוטתיון בגוף או קלוטו, וויטמין D, גם כאן הסוד הוא לא רק לבלוע כדורי קרנוזין, אלא לספק לגוף את אבני הבניין כדי שהתאים (במיוחד תאי השריר ורקמת החיבור) ייצרו ויפרישו אותו בעצמם ישירות למטריצה, לצד עיכוב האנזים שמפרק אותו [מקור1, מקור2, מקור3]:
3. הפעלת מערכת הניקוז הלימפטית וזרימת הנוזלים
הפסולת שפורקה חייבת להתפנות מהאוקיינוס, אחרת היא תשקע ותייצר דלקת (Metaflammation). המערכת הלימפטית היא מערכת הביוב הבלעדית של הנוזל החוץ-תאי [מקור].
- הידרציה דינמית: הגברת שתיית מים טהורים בשילוב חליטות צמחים תומכות מטבוליזם (כמו תה ירוק או חליטות עשירות בפוליפנולים) המסייעות לשטיפת הנוזל הבין-תאי ומשפרות את הפינוי הכלייתי. [מקור1, מקור2, מקור3]
- תנועה מכנית: מערכת הלימפה חסרה משאבה (כמו הלב). היא נעה אך ורק באמצעות התכווצות שרירים. הליכה, עיסוי רקמות עמוק, ותנועה יומיומית חיוניים כדי "לסחוט" את הנוזל המזוהם הישן החוצה ולאפשר לנוזל חדש ונקי להיכנס [מקור1, מקור2 , מקור3].

4. החזרת האיזון האלקטרוליטי וחומציות הרקמה
סוכרת ודלקת כרונית גורמות להחמצה של המטריצה החוץ-תאית, מה שמשבש את התקשורת החשמלית בין התאים.
- עשבי תיבול וירקות עליים ירוקים עשירים בכלורופיל ומינרלים אלקליין (מגנזיום ואשלגן): מסייעים מבחינת רפואת הרשתות וקידומם של מסלולי החלמה להשבת האיזון היוני של הנוזל, ומפחיתים את האותות המכנו-כימיים הדלקתיים שהתאים קולטים מהסביבה המתוחה.
- צירי החלמה – מבוא למחשבה חדשה על בריאות והתחדשות מאת ירון מרגולין
- מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ב', על ההחלמה מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ג', על ההחלמה
- 🔷 המסלולים הביולוגיים שמשקמים את הגוף: p53 — שומר הגנום – חלק ד’ — ירון מרגולין
- חלק ה' – חלבוני ההחלמה: הרשת המולקולרית של הריפוי, ההתחדשות וההומאוסטזיס מאת ירון מרגולין
- חלק ו' על צירי ההחלמה ומה שמפעיל אותם – (signal architecture) מאת ירון מרגולין
- 🔹 מפת מסלולי ההחלמה – 3 צירים – כללי
- המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה – מבט עומק על על 3 המסלולים המרכזיים.
5. גיוס ה"קמיקזות": כך תייצרו את מגן ההקרבה המולקולרי להצלת האוקיינוס התאי
כדי לגייס את לוחמי הקרנוזין (carnosine) למערכה ללא תרופות כימיות, שעלולות לקדם תופעות לואי הרסניות, עלינו להבין את האתגר הביולוגי: הגוף מייצר אנזים בשם קרנוזינאז, שמפרק את הקרנוזין בדם במהירות לפני שהוא מספיק להילחם בסוכר Carnosine metabolism and its degradation by carnosinase. כדי לעקוף את החסימה הזו ולהציף את הנוזל הבין-תאי ב"קמיקזות" שיקריבו את עצמן למען הקולגן, רפואת הרשתות מפעילה אסטרטגיה תלת-ממדית פשוטה וזמינה The anti-glycation effect of carnosine [מקור1, מקור2, מקור3]:
- אספקת חומר הנפץ (בטא-אלנין): הקרנוזין מורכב משתי חומצות אמינו: הקרנוזין מורכב משתי חומצות אמינו: בטא–אלנין והיסטידין [מקור]. כאשר חומצת האמינו בטא–אלנין היא צוואר הבקבוק – בלעדיה התאים אינם יכולים לייצר את הלוחם Beta-alanine supplementation augments muscle carnosine [מקור]. היא נמצאת בשפע במזון אסור לחולי כליות, כמו בשר בקר דל שומן. עבור צמחונים או טבעונים, וחולים שמבקשים לא להיחשף לתופעות לוואי כמו ייצור TMAO בגופם, ושרמות הקרנוזין בגופם נמוכות משמעותית, תיסוף פשוט, נגיש וזול של אבקת בטא–אלנין נקייה פותר את הבעיה ומספק את חומר הגלם לייצור עצמי בתאים Vegetarians have lower muscle carnosine levels. אבקת בטא–אלנין מייצור סינתטי (כימי או ביולוגי): מומלצת בדרך כלל למאמצים עצימים שנמשכים בין חצי דקה למספר דקות, והמינון היומי המקובל הוא לרוב 2 עד 5 גרם. פפריקה, זרעי חילבה (גרגרנית יוונית), זרעי פרג וזרעי חרדל נחשבים לעשירים יחסית בחומצות אמינו שמקדמות סינטזה עצמית בשריר של הבטא-אלנין [מקור1, מקור2] [הערה: זרעי חרדל שלמים או טחונים בלבד. כוספת חרדל (הקליפה, ופנים הזרע – הנבט והאנדוספר) נחשבת למקור חלבון צמחי עשיר מאוד (לרוב סביב 30% עד 40% חלבון), ובה נמצאות חומצות האמינו (כמו היסטידין) שתומכות בייצור של חומרים בגוף.]. הגברת חומצת האמינו היסטידין (Histidine) ממקורות צמחיים: הקרנוזין כאמור לעיל, מורכב משתי חומצות אמינו: בטא-אלנין והיסטידין. למרות שבטא-אלנין היא הגורם המגביל, אספקת היסטידין ממקורות צמחיים איכותיים תבטיח שלגוף יהיו מספיק אבני בניין לייצור עצמי בכבד ובשריר: [מקור1, מקור2, מקור3, מקור4] מקורות צמחיים עשירים בהיסטידין: פולי סויה (טופו, אדממה, נאטו), זרעי המפ, זרעי דלעת, שעועית ועדשים.
- הרדמת הבלם (מעכבי קרנוזינאז טבעיים): כדי למנוע מאנזים הפירוק להשמיד את לוחמי הקרנוזין, אנו משתמשים בפוליפנולים עוצמתיים מהצומח שפועלים כ"משתיקי קול" עבור האנזים ההרסני. צריכה של קקאו טהור וטיפולי, תה ירוק (העשיר ב-EGCG) ותותי עץ שחורים, מעכבת את הפירוק ומאפשרת לקרנוזין לשרוד באוקיינוס החוץ-תאי וללכוד את מולקולות הסוכר הפראיות The anti-glycation effect of carnosine.
- אימוני הפוגות בעצימות גבוהה (HIIT) (אימוני התנגדות קצרים): המפעלים הגדולים ביותר לייצור קרנוזין בגוף הם סיבי השריר המהירים (הסיבים הלבנים). כדי לאלץ אותם להפריש את הקרנוזין ישירות אל רקמת החיבור העוטפת אותם, יש לתת להם אות אבולוציוני של מצב חירום מכני: מחקרים מראים כי אימוני HIIT מגרים את הגוף להעלות באופן עצמאי את רמות הקרנוזין בשרירים, גם ללא כל שינוי בתזונה או נטילת תוספים. מאמץ קצר ועצים גורם לשריר להסתגל לחומציות על ידי ייצור מוגבר של אנזימי סינתזת קרנוזין. כשמדברים על אימוני כוח קצרים הכוונה פעילות נגד התנגדות חזקה או ספרינטים עצימים (HIIT) של 30-60 שניות. אימונים אלו מקפיצים את ריכוז הקרנוזין ברקמות ומקדמים ניקוי של "הבוץ" שנוצר מפגעי הסוכר High-intensity training increases muscle carnosine.
סעיף 6: יתר קורטיזול, חסם הניקוז ונוקשות המטריצה החוץ-תאית (ECM)
כאשר מערכת העצבים הסימפתטית מופעלת ללא הפסקה, עודף הקורטיזול ↑ בנוזל הבין-תאי פועל בשלושה ערוצים הרסניים:
- הרס ארכיטקטורת הקולגן
- חסימת הניקוז הלימפתי
- האצת תהליך הגליקציה (AGEs)
א. המנגנון: כיצד סטרס כרוני מלכלך ומקשיח את המטריצה?
בבואנו לנקות את הנוזל הבין-תאי (הסביבה החוץ-תאית בגופנו), אנו נתקלים לא פעם בחסם עיקש שאינו קשור לתזונה ישירה, אלא למצב הרוחני והמנטלי של המטופל: הורמון הקורטיזול. במצבי סטרס כרוני ↑, חרדה ↑, או שחיקה גופנית מתמשכת ↑, הקורטיזול ↑ הופך מהורמון הישרדותי חיוני לגורם קטבול (מפרק) שמחריב את המטריצה ומקבע בה חסמי פינוי רעלים. "הורמון הסטרס" (הקורטיזול), מבחינה פיזיולוגית הוא הורמון קטבולי (מפרק). כאשר הוא מופרש ברמה גבוהה לאורך זמן ↑, הוא פוגע ישירות בסביבה הבין-תאית (המטריצה החוץ-תאית – ECM):
- פירוק מבני (הרס קולגן): קורטיזול גבוה ↑ מעכב בצורה דרסטית פיברובלסטים (התאים שאחראים על ייצור רקמת החיבור) הוא מעכב ייצור של קולגן ואלסטין ומאיץ את פירוק הקולגן והאלסטין. המטריצה מאבדת מגמישותה והופכת לקשיחה ודחוסה [מקור].
- חסימת הפינוי (הצטברות פסולת): רקמת חיבור נוקשה ודחוסה לוחצת על נימי הלימפה העדינים והמיקרו-כלי דם החוצים אותה. מערכת הניקוז הטבעית "נחנקת", ונוצרת הצטברות מסיבית של פסולת מטבולית, רדיקלים חופשיים ותאים מתים בסביבת התא, מבלי יכולת להתפנות. המטריצה הופכת לדחוסה וקשיחה, "מערכת הניקוז" (מערכת הלימפה והנוזל החוץ-תאי) אינה יכולה לזרום בחופשיות. פסולת מטבולית, תאים מתים ורעלנים נלכדים סביב התאים במקום להתפנות.
- האצת תהליך הגליקציה (AGEs): כחלק ממנגנון "הילחם או ברח", קורטיזול מורה לכבד לשחרר גלוקוז זמין לדם ולנוזל הבין תאי ומעלה, בהמשך הדרך את רמות הסוכר בדם ובנוזל הבין-תאי עולות. סוכר עודף זה נקשר לחלבונים המפורקים במטריצה ומייצר קשרים צולבים (Cross-linking) – תהליך שמקשיח את הרקמה באופן קבוע ומעורר דלקת שקטה (Low-grade inflammation).
ב. הפתרון האדפטוגני: איזון הורמונלי לצורך ניקוי הרקמה
כדי לאפשר ל לחיצות ההחלמה, לזרימת הנוזלים ולתזונה המאזנת לעבוד, חובה להסיר את "פטיש הקורטיזול". מדע האדפטוגנים הצמחיים מציע פתרון סינרגטי, לא סינתטי, המשיב את ההומיאוסטזיס למערכת: פטיש הקורטיזול שמכה בה נח מרגע שנכנסים הצמחים האדפטוגניים, המשחזרים את האיזון (הומיאוסטזיס) ללא חומרים סינתטיים, האהובים כל כך על מתחזקי המחלות ונביאי הרפואה הישנה:
- רודיולה רוזאה (Rhodiola Rosea) – הגנה אקוטית ביום:
השורש הארקטי הקשוח של רודיולה רוזאה (Rhodiola rosea), המכונה "שורש הזהב" פועל כ"בולמי זעזועים" למערכת העצבים [מקור]. נטילתה בשעות הבוקר (לא יותר מ 5 ימים בשבוע ולמשך לא יותר מ 3 חודשים) מונעת מהקורטיזול לזנק בחדות בתגובה לאירועי היום ומגבירה את ייצור האנרגיה התאית (ATP) במטוכונדריה. היא מגנה על הנוזל הבין–תאי מחמצון בזמן שעות הפעילות [מקור1, מקור2, מקור3].- פועלת כמגן מפני זעזועים (Shock absorber). ומונעת מהקורטיזול לזנק בחדות במהלך סטרס יומי או אימון גופני עצים.
- השפעתה על המטריצה: רודיולה רוזאה שומרת על ייצור האנרגיה (ATP) בתאים ומפחיתה את הסטרס החמצוני שפוגע בנוזל הבין-תאי במהלך שעות הפעילות.
- אשווגנדה (Ashwagandha) – שיקום וניקוז בלילה: אשווגנדה: מורידה באופן עקבי ומתמשך את רמות הקורטיזול הכלליות, במיוחד לקראת ערב.
- השפעה על המטריצה: האשווגנדה מאפשרת לגוף להיכנס למצב של מערכת עצבים פאראסימפתטית (מנוחה ועיכול). מערכת הלימפה מתרחבת בזמן הזה ומבצעת את "שטיפת הרקמות" והניקוי העמוק של המטריצה בזמן השינה [מקור1, מקור2 , מקור3].
הורדת רמות הקורטיזול היא תנאי פיזיולוגי הכרחי לריכוך הרקמה, לזרימת נוזלים חופשית ולהחזרת יכולת הריפוי העצמית של הגוף.
ניקוי הנוזל הבין-תאי אינו יכול להתרחש בגוף שנמצא במצב "הילחם או ברח" תמידי. הפחתת קורטיזול באמצעות שילוב אדפטוגנים ממוקדים (רודיולה בבוקר ואשווגנדה בערב) היא תנאי סף פיזיולוגי המאפשר לרקמת החיבור להתרכך, לנוזלים לזרום, ולמנגנוני הניקוי הטבעיים של הגוף לפנות את הפסולת ביעילות [מקור1, מקור2 , מקור3].
הפתרון הרשתי המורחב: כדי לשקם את ה-ECM, הפעלת NRF2 לבדה נדרשת גם בלימת חמצון ואוטופגיה 🧹
הפעלת אוטופגיה (Autophagy 🧹) לפינוי ה-AGEs מחלבוני הרקמה, ואספקת חומרי הזנה ספציפיים (כמו פוליפנולים מזרעי ענבים וליקופן 🍅) לבנייה נקייה [מקור].
שיפוץ ושיחזור מבנה שימור
כשהמטריצה "מזדקנת" או נכון יותר להגדירה נפגעת, ומאבדת גמישות, התאים עצמם ששוחים בתוכה כדגים באוקינוס, מתפקדים פחות טוב — גם אם הם "בריאים" יחסית. לכן טיפול שמנסה לשפר או "לאפס" את הסביבה החוץ-תאית מזווית צרה, כהרגל רפואת הממסד המיושנת, נדרש להפעיל מחדש תהליכי תיקון טבעיים, כפי שהמדע מציע.
(ECM) דורש גישה משולבת המדמה שיפוץ אדריכלי, הכוללת הפעלת נוגדי חמצון דרך מסלול ה-NRF2, ניקוי תוצרי גליקציה (AGEs) באמצעות אוטופגיה 🧹, ושימוש ברכיבים תזונתיים לחידוש סיבי הקולגן. אסטרטגיה זו הופכת את רקמת החיבור למבנה גמיש, המגן מפני פיברוזיס וזקנה מערכתית [מקור1, מקור2 , מקור3, מקור4].
מדוע NRF2 וסולפוראפן (נבטי ברוקולי) הם רק ההתחלה?
כשכתבתי על נבטי ברוקולי 🥦 ו-NRF2 בעבר, נגעתי במנגנון הגנה קריטי. NRF2 הוא מאסטר-רגולטור נוגד חמצון שמפחית את העקה החמצונית ב-ECM ובולם את הפעלת ה-MMPs ההרסניים [מקור1, מקור2].
אבל ברפואת רשתות עמוקה, הפעלת יתר של NRF2 לבדו ברקמת החיבור עלולה דווקא לעכב ייצור קולגן חדש (כמנגנון הגנה מהצטלקות/פיברוזיס). לכן, כדי לבצע ביוהאקינג אמיתי לרקמת החיבור, אנחנו צריכים לשלב כוחות ברשת: [מקור]
- מצד אחד (בלימה): לעורר את NRF2 🥦 כדי לעצור חמצון והרס.
- 🧹מצד שני (פינוי): להפעיל את האוטופגיה (Autophagy) כדי שתאי הפיברובלאסט יבלעו ויפרקו את חלבוני ה-ECM הפגומים והמסוכרים (AGEs).
- מצד שלישי (בנייה): לספק חומרי הזנה ספציפיים (כמו פוליפנולים מזרעי ענבים 🍇ם וליקופן 🍅) שמגינים על סיבי האלסטין ומאפשרים בנייה תקינה, נקייה וגמישה של המטריצה [מקור1, מקור2, מקור3, מקור4, מקור5].
לסיכום
רקמת החיבור והמטריצה החוץ-תאית (ECM)
המטריצה החוץ-תאית (ECM) אינה רק "פיגום" פסיבי התומך בתאים. היא האוקיינוס הפנימי שבו כל תא בגופנו חי ושוחה. המדע מגדיר אותה כיום כאחד מסימני ההיכר הרשמיים של ההזדקנות (Hallmarks of Aging).
כמו מפרץ צלול או אולם קונצרטים, כל עוד המטריצה גמישה ונקייה – התאים מקבלים חמצן, מזון ואותות תקינים. אך כאשר היא מתקשחת, מתלכלכת ומאבדת גמישות, התאים מתחילים להגיב כאילו הם תחת מתקפה מתמדת. התוצאה: דלקת כרונית, ירידה בתפקוד והאצה של תהליכי הזדקנות.
שלושת האויבים העיקריים של המטריצה:
- תוצרי גליקציה מתקדמים (AGEs) – סוכר עודף יוצר קשרים צולבים בקולגן ובאלסטין, הופך את הרקמה לקשיחה ומאותת לתאים "מצב חירום".
- אנזימי MMPs – במצב של דלקת כרונית הם יוצאים מאיזון ומפרקים את הרקמה ביתר.
- שברי חלבון רעילים – תוצרי הפירוק עצמם הופכים למקור נוסף של דלקת מערכתית (Inflammaging).
הפתרון הרשתי המשולב מציע לא לתקן רק את התא, אלא לנקות ולשקם את הסביבה שבה הוא חי:
- הפעלת אוטופגיה לפינוי חלבונים פגומים ו-AGEs.
- שימוש במעכבי גליקציה (בעיקר קרנוזין וחומצה אלפא-ליפואית) ובאבני הבניין שלהם (בטא-אלנין והיסטידין).
- שיפור הניקוז הלימפטי באמצעות תנועה, הידרציה וחליטות.
- איזון אלקטרוליטי והפחתת חומציות הרקמה.
- הורדת קורטיזול כרוני באמצעות אדפטוגנים (רודיולה בבוקר, אשווגנדה בערב).
- שילוב של NRF2 (נבטי ברוקולי / סולפוראפן) עם חומרי בנייה כמו פוליפנולים מזרעי ענבים וליקופן.
המסר המרכזי: הזדקנות הרקמות אינה מתחילה רק בתוך התא. היא מתחילה כאשר האוקיינוס שסביבו משתנה. שיקום המטריצה החוץ-תאית הוא אחד המפתחות החשובים להחזרת גמישות, תפקוד ואריכות ימים אמיתית.
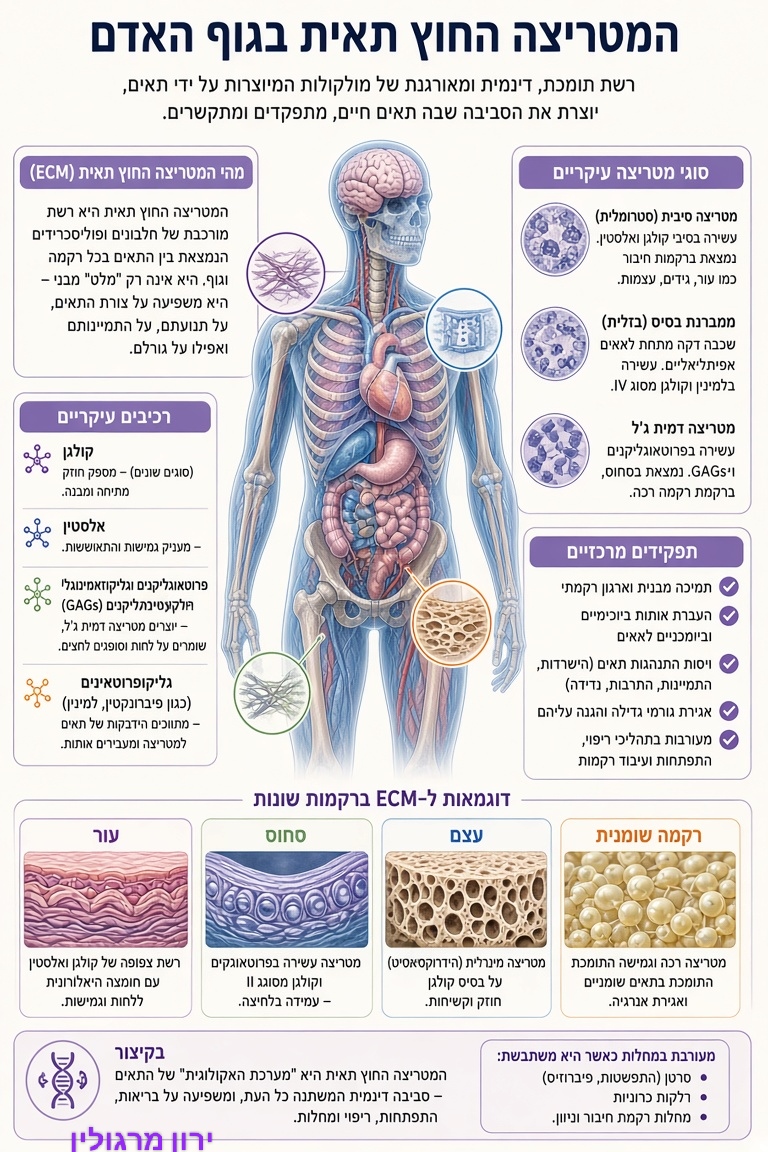
המטריצה החוץ-תאית (ECM) היא האוקיינוס הפנימי שבו חיים כל תאי הגוף. כשהיא גמישה ונקייה – התאים מתפקדים היטב. כשהיא מתקשחת ומתלכלכת (בעיקר בגלל AGEs מסוכר עודף, דלקת וקורטיזול גבוה) – התאים נכנסים למצב דלקתי כרוני והזדקנות מואצת.
הפתרון אינו רק "לתקן את התא", אלא לנקות ולשקם את הסביבה שלו: הפעלת אוטופגיה, עיכוב יצירת AGEs (קרנוזין + חומצה אלפא-ליפואית), שיפור ניקוז לימפטי, הורדת קורטיזול (אדפטוגנים) והפעלת NRF2 יחד עם חומרי בנייה.
שיקום המטריצה הוא מפתח מרכזי לגמישות, תפקוד ואריכות ימים.
ירון מרגולין
נשארו לך שאלות
🔬אשמח להשיב על כל שאלה
בבקשה לא להתקשר משום שזה פשוט לא מאפשר לי לעבוד – אנא השתמשו באמצעים שלפניכם
למען הסר ספק, חובת התייעצות עם רופא (המכיר לפרטים את מצבו הבריאותי הכללי של כל מטופל או שלך) לפני שימוש בכל תכשיר, מאכל, תמצית או ביצוע כל תרגיל. ירון מרגולין הוא רקדן ומבית המחול שלו בירושלים פרצה התורה כאשר נחשפה שיטת המחול שלו כבעלת יכולת מדהימה, באמצע שנות ה – 80 לרפא סרטן. המידע באתר של ירון מרגולין או באתר "לחיצות ההחלמה" (בפיסבוק או MARGOLINMETHOD.COM ), במאמר הנ"ל ובמאמרים של ירון מרגולין הם חומר למחשבה – פילוסופיה לא המלצה ולא הנחייה לציבור להשתמש או לחדול מלהשתמש בתרופות – אין במידע באתר זה או בכל אחד מהמאמרים תחליף להיוועצות עם מומחה מוכר המכיר לפרטים את מצבו הבריאותי הכללי שלך ושל משפחתך. מומלץ תמיד להתייעץ עם רופא מוסמך או רוקח בכל הנוגע בכאב, הרגשה רעה או למטרות ואופן השימוש, במזונות, משחות, תמציות ואפילו בתרגילים, או בתכשירים אחרים שנזכרים כאן.
physician (who knows in detail the general health of each patient or yours) before using any medicine, food, extract or any exercise. The information on Yaron Margolin's website or the "Healing Presses" website (on Facebook or YARONMARGOLIN.COM), in the above article and in Yaron Margolin's articles are material for thought – philosophy neither recommendation nor public guidance to use or cease to use drugs – no information on this site or anyone You should always consult with a qualified physician or pharmacist regarding pain, bad feeling, or goals and how to use foods, ointments, extracts and even exercises, or other remedies that are mentioned as such
מאמרים אחרונים
- רקמת החיבור והמטריצה החוץ-תאית (The Extracellular Matrix – ECM)
- צירי החלמה – מבוא למחשבה חדשה על בריאות והתחדשות מאת ירון מרגולין
- מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ב', על ההחלמה מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ג', על ההחלמה
- 🔷 המסלולים הביולוגיים שמשקמים את הגוף: p53 — שומר הגנום – חלק ד’ — ירון מרגולין
- מנגנוני תיקון פנימיים בגוף: מודל אינטגרטיבי להבנת תפקוד הכליות והמערכת כולה – 3 צירי ההחלמה המרכזיים
- המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה
- מיומטי תנועה – התקווה החדשה לחולי כליות: תרכובות שמחקות את צירי ההחלמה של פעילות גופנית
- סיפורם המרתק של בני שבט הינאומאמי והסוד הפיזיולוגי המדהים של כליות הגמל, שמסביר כיצד מערכת הישרדות פראית הופכת לקטלנית בעולם המודרני נוכח הארוע האבולוציוני של אי-ספיקת לב ותפקוד הכליות/ ירון מרגולין
- הכליות הן הרופא הפנימי שאיבד את המפתחות בעת חולי: מסע מולקולרי אל סודה של רשת חלבוני החלמת הכליות ומסלול האיתות TGF–β אשר משנה את עורו שוכח את תפקידו כרופא וחוצה את הקווים.
- שומרי האוצר הזעיר – רפואת העתיד – חלק א' / ירון מרגולין
- שומרי האוצר הזעיר – חלק ב' רפואת העתיד / ירון
- על הגאונות והגאון
- 📘 The Biological Pathways That Restore the Body: A New Perspective on Recovery
- הגוף יודע על המחלה: כיצד לזהות אותה באמצעות מבט עומק בבדיקות המעבדה ובתסמינים הגופניים
- דיאליזה: מאבק הירואי שאינו ריפוי
- רעלים אורמיים מקדמים דיאליזה – הוכח לאחרונה שרעלנים אורמיים קשורים למיקרוביוטה של המעי הגס – איך להפעיל צירי החלמה ביולוגיים רדומים שנמצאים בגוף ללא תרופות ולהימנע מדיאליזה.
- מכתב תודה ממחלימת כליות בתוך ארבעה חודשים.
- מסלול איתות Wnt
- נתיבי החלמה: הסיפור הסמוי של הרשת המטבולית בגוף האדם ומלכודת התרופות – ירון מרגולין
- רשתות של הידרדרות ביולוגית בגוף האדם, האם גם רשתות החלמה פועלות בגופנו❓ ירון מרגולין
- הדור הבא של הרפואה המותאמת אישית: חיישני זיעה עשויים בעתיד לספק משוב ביולוגי רציף, שיסייע להתאים תזונה, אימון והתאוששות לכל אדם – מחקר חדש / ירון מרגולין
- שבר דו־גדילי ב־DNA: מסלולי התיקון, בחירת התא בין HR ל־NHEJ, והשלכות על סרטן, הזדקנות ומחלת הכליות, חלק א' / ירון מרגולין
- שבר דו־גדילי ב־DNA: מסלולי התיקון, בחירת התא בין HR ל־NHEJ, והשלכות על סרטן, הזדקנות ומחלת הכליות / חלק ג' על ההחלמה ומה ניתן לעשות❓
- להחלים מלוקמיה פרומיאלוציטית חריפה בילדים – האם ויטמין A משתתף בתהליך ההחלמה❓- מחקר חדש ותוצאות מרשימות
- כבד שומני ❓- הכבד כמראה מערכתית: צירי מחלה, מול צירי החלמה – מבט אל רפואת העתיד
- סוויטה לירית" (Lyric Suite) אנא סוקולוב
- גמישות העורקים – רפואת העתיד – חלק א'
- למה חולים שנוטלים כדורים להפחתת לחץ דם גבוה, נדרשים בדחיפות להפעיל צירי החלמה ביולוגיים רדומים שנמצאים בגוף כדי לעכב דיאליזה
- איך לא להגיע לדיאליזה
- תובנות חדשות אין להתחיל דיאליזה לפני בדיקת – מצבו של ציר FGF23-Klotho
- פחות מתים – טכניקת דיאליזה חדשה שבולמת את קצב המתים ומגבירה את ההישרדות בכשל כליות.
- איך התפתחו לחיצות ההחלמה בבית מחול ירון מרגולין בירושלים
- המזון הבסיסי
- המזון הבסיסי והחומצי – מה כן ומה לא נכון. מה עובד על ההחלמה ומה הן אגדות? חומציות מגיעה מתזונה והיא קמה והורסת את הכליות
- קפה – הרבה יותר מקפאין. אבל מה לגבי חולי כליה?
- תובנות חדשות אין להתחיל דיאליזה לפני בדיקת – מצבו של ציר FGF23-Klotho
- עיגול פינות מקשה על החלמת הכליות. מיומנו של מאסטר בהחלמת הכליות – הכליות לא סולחות על הזנחה:
- תובנות חדשות אי-ספיקת כליות תלויה במצבו של ציר FGF23-Klotho
- דיאליזה: מאבק הירואי שאינו ריפוי
- פחות מתים – טכניקת דיאליזה חדשה שבולמת את קצב המתים ומגבירה את ההישרדות בכשל כליות.
- שכיחות של דיכאון וחרדה בקרב חולי כליות
- המיקרוביום הכלייתי (האורוביום Urobiome) נחשף
- ⭐ התחדשות כלייתית: מקרה נדיר של ריפוי טבעי — ומה שהוא מלמד את כולנו
- על ההחלמה – למה הגוף לא מחלים — וכיצד להחזיר לו את היכולת הטבעית לעשות זאת
- שינה חשובה להחלמה – 🌜זמן הריפוי החשוב מתרחש בלילה
- ניקוי כבד טבעי – הדרך הפשוטה לבריאות מתחדשת
- מיומטי תנועה – התקווה החדשה לחולי כליות: תרכובות שמחקות את צירי ההחלמה של פעילות גופנית
- הגוף יודע על המחלה: כיצד לזהות אותה באמצעות מבט עומק בבדיקות המעבדה ובתסמינים הגופניים
- איך התפתחו לחיצות ההחלמה בבית מחול ירון מרגולין בירושלים
- המזון הבסיסי והחומצי – מה כן ומה לא נכון. מה עובד על ההחלמה ומה הן אגדות? חומציות מגיעה מתזונה והיא קמה והורסת את הכליות
- הביוכימיה של ההחלמה: מפת העל של החיים
- השמנה בטנית – סטרס ביולוגי — מנגנון העל שמסביר השמנה בטנית גם בלי אכילת יתר. למה לא יורדים בבטן בתהליכי הרזיה מסודרים?
- על הגאונות והגאון
- מנגנוני תיקון פנימיים בגוף: מודל אינטגרטיבי להבנת תפקוד הכליות והמערכת כולה
- סמנים ביולוגיים מוקדמים – ביומרקרים חדשים – ככל האפשר לנפגעי הכליה בבדיקות דם עבור מחלת כליות חריפה AKI או אי ספיקת כליות CKD יכולה להציל את חייהם של אלפי אנשים.
- עשרת המזונות הבריאים ביותר לחולי כלייה – על השעורה (דגן) ופעולות נוגדות דלקת כולל עיכוב גורם נמק גידול אלפא – Tumor necrosis factor alpha (TNF-α).
- להפעיל מסלולי החלמה ביולוגיים בעת מחלה כרונית קשה עכשיו – היו סבלניים.
- הקשר בין כליה לבריחת סידן – תפקוד לא מוערך של הכליה במניעת אוסטאופורוזיס
- בשר, כליות וזרימת דם מואצת: כיצד תזונה מהחי פוגעת במערכת הסינון של הגוף
- השפעה קלינית של ריבס על הטיפול באי ספיקת כליות כרונית: כפי שמשתקף במחקרים.
- טסטוסטרון אינו גורם, מזרז או מחמיר סרטן ערמונית – וה-FDA מתחיל להודות בזה
- חימצון האינסולין והגברה של חומצת שתן בדם, גאוט, אי ספיקת כליות, שבץ לב, סוכרת ודלקת חריפה "בלתי מוסברת"
- רעלים אורמיים מקדמים דיאליזה – הוכח לאחרונה שרעלנים אורמיים קשורים למיקרוביוטה של המעי הגס – איך לקדם את יכולת הגוף להפעיל חלבוני החלמה ללא תרופות ולהימנע מדיאליזה.
- טסטוסטרון אינו גורם, מזרז או מחמיר סרטן ערמונית – וה-FDA מתחיל להודות בזה
- חימצון האינסולין והגברה של חומצת שתן בדם, גאוט, אי ספיקת כליות, שבץ לב, סוכרת ודלקת חריפה "בלתי מוסברת"
- "אילוף הליות הסוררות''. מאמר היסוד על דרכי הפעלת מסלולי החלמה בגוף לקדם שיקום של הכליות.
- באיזה אופן תהליך המתילציה משפיע על ההחלמה ממחלות כרוניות כמו מחלת הכליות
- ירידה בתפקוד הכליות – מה לעשות?
- בשר, כליות וזרימת דם מואצת: כיצד תזונה מהחי פוגעת במערכת הסינון של הגוף
- מדד חדש לבריאותך – ביטול האסרוסקלרוזים
- אי ספיקת כליות – טיפול ללא תרופות
- הקורטיזול וכאבים בבית החזה וביד שמאל
- אוראה – למה האוראה מוגברת ומופחת בדם Urea
- להוריד קריאטינין, אוריאה ולקדם הפעלה של צירי החלמה ביולוגיים בגוף ללא תרופות מאי ספיקת כליות
- ההומוציסטאין – האויב הגדול של האנדותל ובריאות העורקים
- אריתרופויטין (EPO) Erythropoietin
- התרופות והרע – ויטמין D3
- על בדיקות הדם
- אנמיה למה חסר לי ברזל?
- רוצה להפעיל מנגנוני שיקום טבעיים בגוף בעת פגיעה במערכת העיכול ללא תרופות?
- כשהמוסר מתנתק מן החיים: כיצד הפכה האידאולוגיה הנאורה מביקורת מוסרית לעמדה הרסנית ומשתקת
- המוח נלחם על חייו. p62, אוטופאגיה והמשמעות החדשה של אלצהיימר
- המיקרוביום הכלייתי (האורוביום Urobiome) נחשף
- תאי T בזמן החלמה: כיצד מערכת החיסון מתכיילת בלילה כדי לרפא ולא רק להילחם בגורמים למחלה
- מבוא – גוף, רגש ומהפכות נסתרות בתולדות המחול
- גלוטתיון – מנוע התיקון המרכזי של הגוף
- למה חולים שנוטלים כדורים להפחתת לחץ דם גבוה, מסתכנים יותר בקידום הדיאליזה
- סמנים ביולוגיים מוקדמים – ביומרקרים חדשים – ככל האפשר לנפגעי הכליה בבדיקות דם עבור מחלת כליות חריפה AKI או אי ספיקת כליות CKD יכולה להציל את חייהם של אלפי אנשים.
- חימצון האינסולין והגברה של חומצת שתן בדם, גאוט, אי ספיקת כליות, שבץ לב, סוכרת ודלקת חריפה "בלתי מוסברת"
- בפשטות לקדם חלבוני החלמה שנמצאים בגוף האדם במחלת ריאות חסימתית כרונית
- רוצה להפעיל מסלולים ביולוגיים שמקדמים תפקוד תקין בגוף בסוכרת
- שחיקת סחוס, למה לסבול? – ללא ניתוח ללא תרופות – טיפול להחלמה
- עורקים גמישים – הסוד והדרך לזכייה בבריאות מחדש
- ההומוציסטאין – האויב הגדול של האנדותל ובריאות העורקים
- The Biological Pathways That Restore the Body: A New Perspective on Recovery
- אילוף הכליות הסוררות
- Kidney FailureTaming the Rogue KidneysVital Information
- Learn the 8 Principles That Revive Kidney Function
- Reconstruction of erythropoietin by the kidneys without drugs
- Six degrees of guilt – Why do people avoid living their dreams?
- Of sexual desire and the wish for intimacy and the two different languages in which they materialise:
- מוכרחים להתחיל בדיאליזה טרם נבחנה רמתו של גורם צמיחה פיברובלסט 23
- The day will come when we will sit in front of the fireplace
- מכתב תודה ממחלימת כליות בתוך ארבעה חודשים
- ירידה בתפקוד הכליות – מה לעשות?
- אי ספיקת כליות – צום חלבונים
- צום חי – הוא תרופה טבעית. אזהרה לקטונים (Lactones) –
- להחזיר את השליטה לכליה: תזונה נגד ספיגת היתר של הסוכר – איך לאזן את החלבון SGLT2
- תפקוד הכליה בשעות הלילה: השפעת המקצב הצירקדי על הפילטר הביולוגי
- טעם מר מעודד הפרשת רעלים מהכליות – רשימת צמחים בעלי טעם מר מאת ירון מרגולין
- הלשון של הכליות
- כיצד הכליה מטפלת בחלבונים
- אינסולין ותפקיד הכליות במטבוליזם ופינוי
- אינסולין ותפקיד הכליות במטבוליזם ופינוי
- עוצמתם של נבטי הברוקולי כתרופה לחולי כליה
- תפקוד הכליה כבלוטה הורמונלית: סקירה מדעית
- דלקת כליות אינטרסטיציאלית – השתמ
- פרנצ'סקו מסינה Francesco Messina אלהים באדם שחיפש אחר היופי
- ויטמין השמש ויטמין D-
- רוצה להפעיל צירי החלמה ביולוגיים בגוף ביתר לחץ דם?
- ראיית המעמקים – כניסה לטרקלין או על החיים האמתיים.
- התודעה השלילית
- ביקורת מבזה
- על לחם חוקם של העסקים פרק א
- על לחם חוקם של העסקים פרק ב
- על לחם חוקם של העסקים פרק ג
- על לחם חוקם של העסקים פרק ד
- 🇯🇵 腎機能と全身の健康に関わる生体調節メカニズムの概要
- 🇬🇧 Understanding Intrinsic Regulatory Mechanisms:
- על הגאונות והגאון
- 🇯🇵 腎機能と全身の健康に関わる生体調節メカニズムの概要
- 🇬🇧 Understanding Intrinsic Regulatory Mechanisms:
- 🇩🇪 Körpereigene Regulationsmechanismen verstehen: Bedeutung für Nieren, Herz, Gehirn und weitere Organsysteme
- 📘 Карта путей восстановления
- Mapa de las Vías de Recuperación
רקמת החיבור והמטריצה החוץ-תאית (The Extracellular Matrix – ECM)
כשהמטריצה "מזדקנת" או נפגעת, התאים עצמם מתפקדים פחות טוב — גם אם הם "בריאים" יחסית. לכן טיפול שמנסה לשפר או "לאפס" את הסביבה החוץ-תאית של גוף האדם יכול, להפעיל מחדש תהליכי תיקון טבעיים ולקדם שיקום, ניקוי ובניה מחדש. עדויות פרה-קליניות טובות מלמדות: ECM צעיר יכול "להצעיר" תאי גזע מבוגרים, חומרים שמעכבים צילוב-יתר (כמו מעכבי LOX) או משנים את הקשיחות של המטריצה משפרים תפקוד על פי מחקרים חדשים. ירון מרגולין




