
חלק ב' בעמוד זה למטה,
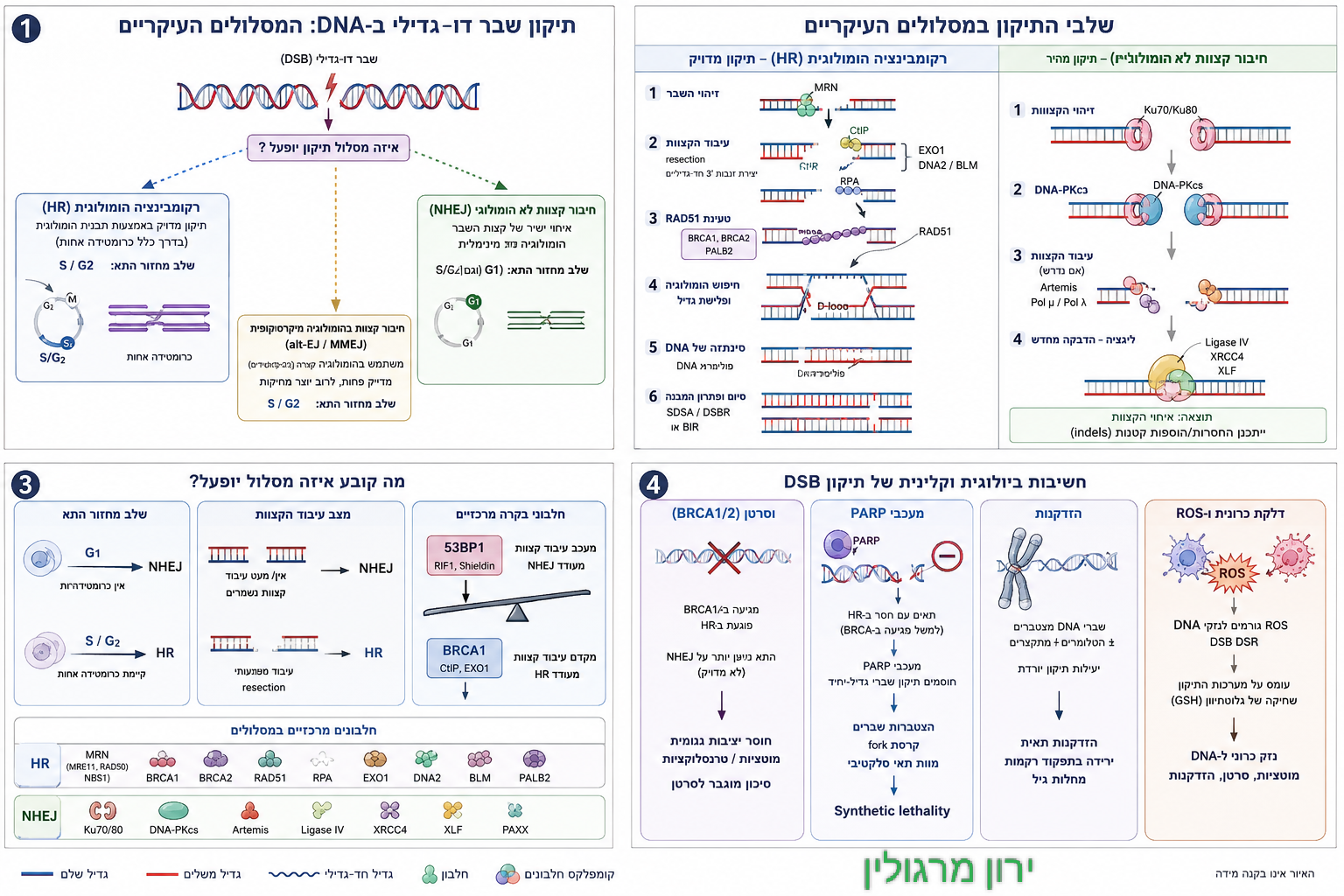
שני המסלולים — רקומבינציה הומולוגית (HR) ו־חיבור קצוות לא הומולוגי (NHEJ) — הם שני צירי ההצלה המרכזיים של התא כאשר נוצר שבר דו־גדילי ב־DNA (Double-Strand Break, DSB).
זה אחד הנושאים היפים בביולוגיה של התא. מהלך שמראה איך הגוף בוחר בין דיוק לבין מהירות, בין שחזור נאמן לבין הישרדות מיידית.
ירון מרגולין
מהם שני הצירים הללו – המנגנונים?
כאשר נגרם שבר דו–גדילי ב-DNA (הנזק החמור ביותר שיכול לקרות לגרעין התא), לתא יש שתי דרכים עיקריות לטפל בזה:
- NHEJ (Non-Homologous End Joining): מנגנון "עזרה ראשונה" מהיר ומחוספס. הוא פשוט מדביק את שני הקצוות השבורים בחזרה זה לזה. המנגנון פעיל לאורך כל מחזור התא, אך הוא נוטה לטעויות (Error-prone) ולעיתים קרובות גורם להוספה או מחיקה של בסיסים (מוטציות).
- HR (Homologous Recombination): מנגנון תיקון מדויק לחלוטין (Error-free). הוא משתמש בכרומטידה האחות (עותק זהה של ה-DNA) כתבנית כדי להעתיק ממנה את המידע החסר ולתקן את השבר בצורה מושלמת. הוא פעיל רק בשלבי מחזור התא שבהם ה-DNA כבר עבר הכפלה (שלבי S ו-G2).
פתיח
בכל תא חי טמון פרדוקס: כדי להמשיך להתקיים, עליו לשמור בקפדנות על יציבות החומר הגנטי, אך באותה שעה הוא חשוף ללא הרף לפגיעות המאיימות לקרוע אותו מבפנים. אחת הפגיעות החמורות ביותר היא שבר דו־גדילי ב־DNA (Double-Strand Break, DSB) — מצב שבו שני גדילי ה־DNA נחתכים, והמידע הגנטי עצמו עומד בסכנת אובדן, עיוות או חיבור שגוי. כאשר נזק כזה אינו מתוקן כראוי, התא עלול לאבד את יכולתו להתחלק, להיכנס להזדקנות תאית, למות, או לחלופין לצבור מוטציות מסוכנות ואי־יציבות כרומוזומלית.
לשם התמודדות עם איום זה פיתח התא שני מסלולי תיקון עיקריים: רקומבינציה הומולוגית (HR) — מסלול מדויק יחסית המשתמש בכרומטידה האחות כתבנית לתיקון, ו־חיבור קצוות לא הומולוגי (NHEJ) — מסלול מהיר יותר, המאפשר איחוי של קצות DNA שבורים גם ללא תבנית מלאה. הבחירה בין המסלולים ⚖️ אינה טכנית בלבד; היא משקפת הכרעה עמוקה על גורל התא: האם יתוקן במדויק🛠️, ישרוד במחיר של טעות, ייכנס לעצירת מחזור, או ייסחף למסלול של דלקת, פיברוזיס והידרדרות תפקודית.
משמעות הדברים בולטת במיוחד במחלת הכליות. הכליה היא איבר עתיר פעילות מטבולית, החשוף לעקה חמצונית, לדלקת כרונית, לאיסכמיה, לרעלנים אורמיים ולעומס מתמשך על המיטוכונדריה. תנאים אלו עלולים להגביר פגיעות ב־DNA ולהעמיס על מערכות התיקון. לכן, במחלת כליות כרונית אין לראות בנזקי DNA רק סיכון מרוחק להתמרה סרטנית, אלא גם מנגנון מרכזי היכול לדחוף תאים כלייתיים אל הזדקנות, אובדן יכולת התחדשות, הפרשת אותות פרו־פיברוטיים וצלקת מתקדמת של הרקמה.
מאמר זה מבקש להתבונן בשבר הדו־גדילי (DSB) לא רק כאירוע מולקולרי נקודתי, אלא כצומת הכרעה ביולוגי: נקודה שבה מוכרע אם התא והרקמה ינועו לעבר תיקון והתאוששות — או לעבר פיברוזיס, אובדן נפרונים, הזדקנות ולעיתים גם סרטן. מתוך כך נבחן את שני מסלולי התיקון המרכזיים, את מנגנוני הבקרה המכוונים אותם, ואת האופן שבו כשל בתיקון DNA עשוי להשתקף בלב מחלת הכליות עצמה.
קריאה נעימה.
איור: ציר הזמן של מחזור התא והמחשה לגבי הזמן המדויק של מנגנון ה-HR נכנס לפעולה:
תהליך הכפלת ה-DNA 🧬
│
┌───────────────────────┴───────────────────────┐
▼ ▼
[ שלב G1 ] ───► ───► [ שלב S ] ───► ───► [ שלב G2 ] ───► [ שלב M ]
(צמיחת התא) (שכפול ה-DNA) (בקרת איכות) (חלוקה)
│ │ │
│ ├───────────────────────┤
│ │ │
▼ ▼ ▼
┌──────────────┐ ┌────────────────────────────────────────────────┐
│ NHEJ בלבד │ │ פעילות מנגנון HR │
│ │ │ (יש כרומטידה אחות זהה המשמשת כתבנית לתיקון) │
└──────────────┘ └────────────────────────────────────────────────┘
לפני שניכנס לעומקם של הדברים 📊 הסבר קצר על זמני הפעילות:
- שלב G1 (לפני הכפלה): לתא יש רק עותק אחד של ה-DNA. מכיוון שאין "גיבוי" שממנו אפשר להעתיק מידע, מנגנון ה-HR אינו מסוגל לפעול. בשלב זה התא מסתמך רק על NHEJ המהיר והפחות מדויק.
- שלבי S ו-G2 (תוך כדי ואחרי ההכפלה): ה-DNA משוכפל וישנן שתי כרומטידות אחיות זהות לחלוטין. זהו תנאי הסף להפעלת ה-HR – כעת האנזימים יכולים לקחת את הגדיל השבור, להשוות אותו לקופי המדויק שנמצא ממש לידו, ולתקן את הנזק ללא שום טעויות.
p53: השופט והמוציא לפועל של התא הפגוע
בחלק ד' – על שומר החומר הגנטי🔹🐦🔥 P53 🐦🔥 והנחיות טבעיות של הגוף ל: "החלמה" אני מציג את החלבון המגן ועל דרכים להפעלתו לקידומם של תהליכי ההחלמה.
כאשר נגרם שבר דו–גדילי (DSB), התא אינו יכול להרשות לעצמו להמשיך "באורח חייו הרגיל" – חלוקה של תא עם DNA שבור תוביל למותו או להפיכתו לתא חולה בדרך כלל לתא סרטני. כדי למנוע זאת, התא מפעיל מערכת איתור ואזעקה משוכללת ביותר היא ה- DDR (DNA Damage Response).
חלבון ה-p53, המכונה "שומר הגנום", הוא המפקד העליון של המערכת הזו. להלן שלבי האיתור, האיתות וההחלטה של התא:

האיור ממחיש את גוף האדם כמפה חיה של מסלולי התחדשות ושיקום.
הדמות של אלכסנדר הגדול לוחם שיוצא למסע גדול – מסמלת את האדם הנועז שיוצא מתקופת נזק והצטברות אל עבר ריפוי והתחדשות. מסע של מעברים מרפואת התחזוקה להפעלתם של מסלול החלמה. רפואת העתיד.
קווי האור הזוהרים באיור מסמנים את הצירים הביולוגיים המרכזיים: p53 Activation, AMPK Pathway, Autophagy, SIRT1, Nrf2, Mitochondrial Biogenesis, DNA Repair ועוד. מחוץ לגוף מצוינים חומרי ההפעלה הטבעיים שיכולים לתמוך בכל מסלול. כמו "גיבורי השומר", לוחמי העל שהגנו והובילו לגאולת הארץ השקועה בביצה, ביניהם סבא שלום מרגולין, אלכסנדר זיד וטרומפלדור – כך גם גופנו יכול להפוך ביצה מיושנת וחולה, מערכת שקועה ברעלים, לעצירת הזיהום ומקור הרעל, נוכח אתגר הכולל מתח קל וידע רחב להזדמנות של תיקון עמוק והתחדשות מלאה.
מנהל מערכת התיקון: כיצד p53 מנווט את הטיפול בשברי DNA
🚨 שלב 1: הזיהוי והאזעקה המיידית (ATM ו-γH2AX)
ברגע שמתרחש ה-DSB, קצוות ה-DNA הקרועים נחשפים. חלבונים ייעודיים נקשרים אליהם מיד ומגייסים אנזים מפתח בשם ATM (אנזים שתפקידו לאותת על שברים דו–גדיליים).
- סימון אזור האסון: אנזים ה-ATM מזרחן (מוסיף קבוצת פוספט) לחלבון המבנה של ה-DNA שנקרא H2AX. הגרסה המזורחנת שלו נקראת γH2AX (גמא-H2AX).
- חלבוני γH2AX יוצרים מעין "פנסים מולקולריים" זוהרים סביב השבר. האיתות המהיר הזה קורא לכל אנזימי התיקון (כמו אלו של HR ו-NHEJ) להגיע לנקודה.
📣 שלב 2: הפעלת ה-p53 (העברת פקודת החירום)
במצב שגרה, חלבון ה-p53 מיוצר ומפורק מיד (על ידי חלבון בשם MDM2) כדי שלא יפריע לתא. הוא נשמר ברמות נמוכות מאוד.
- הפרדת המפרק: כאשר ה-ATM מופעל בגלל ה-DSB, הוא מזרחן את p53 ואת המפרק שלו (MDM2). הזרחון הזה גורם להם להיפרד.
- ייצוב ה-p53: כעת, ה-p53 יציב, מוגן מפני פירוק, ורמותיו בתא קופצות בבת אחת. הוא נודד ישירות לגרעין התא ופועל כגורם שעתוק (מפעיל של גנים אחרים).
🛑 שלב 3: עצירת מחזור התא (עצירת חירום ב-G1 או G2)
הפעולה הראשונה של p53 היא לעצור את חלוקת התא כדי לתת זמן לתיקון:
- ה-p53 מפעיל גן שמייצר חלבון בשם p21.
- חלבון ה-p21 הוא ה"ברקס" של התא – הוא חוסם את חלבוני ה-CDK (אלו שדוחפים את התא לעבור משלב לשלב במחזור התא).
- התא נעצר (Cell Cycle Arrest), בדרך כלל בנקודת הביקורת של G1/S או G2/M. כעת התא "קפוא" בזמן, והאנזימים יכולים להתחיל לעבוד על ה-DSB (בין אם בדרך של NHEJ המהירה או בדרך של HR המדויקת בשלבים המאוחרים).
⚖️ שלב 4: גזר הדין – תיקון או התאבדות (Apoptosis)
החלק המרתק ביותר ב-p53 הוא יכולת קבלת ההחלטות שלו. הוא משקלל את כמות האיתותים של ה-DSB ומחליט על גורל התא:
- אם הנזק קטן ובר-תיקון: ה-p53 משאיר את הברקס (p21) לחוץ, ומפעיל גנים שמעודדים את מערכות התיקון (כמו חלבוני ה-HR). ברגע שהנזק מתוקן, האיתות מ-γH2AX נפסק, ה-p53 מתפרק, והתא חוזר למחזור החיים שלו.
- אם הנזק קטסטרופלי (יותר מדי DSBs): ה-p53 מבין שהסיכון ליצירת תא סרטני גבוה מדי. הוא עובר להפעיל גנים קטלניים (כמו Bax ו-Puma) שיוצרים חורים במיטוכונדריה. התוצאה היא אפופטוזיס (Apoptosis) – מוות תאי מתוכנן ונקי, שמקריב את התא הבודד למען בריאות האורגניזם כולו.
💡 נקודה למחשבה האם מקורות הנזק בצלחת שלך
כאשר חומרים עליהם ארחיב בפרק בהמשך ביניהם חומרים שהגוף מיצר בשל מחסור בחומרי הזנה ואבני בניין כמו יתר הומוציסטאין, TMAO או סטרס כרוני (קורטיזול) מייצרים DSBs ללא הפסקה, ה-p53 מופעל שוב ושוב. לאורך זמן, הדבר מוביל להתרוקנות של מאגרי תאים בריאים (בגלל אפופטוזיס מוגבר) או לאבדן תפקוד תאי, נושא שמכונה במחקר הזדקנות מואצת של הרקמה (Cellular Senescence), וזו הסיבה המולקולרית לכך שהנמצא בצלחת שלך קריטי ומקור מרכזי לסטרס תאי. תזונה לקויה מקדמת מחלות כרוניות אבדן תפקוד מה שמאפשר לחשוף בפניך את עולם התרופות, אבל גם הן לפעמים יוצרות סטרס תאי, כך שנפתח מעגל מורכב ומאוד מסובך שמנת מתאימה 🥗, יכלה למנוע..
אפשר כמובן לבדוק מה קורה כאשר ה-p53 עצמו נפגע או עובר מוטציה (כפי שקורה במחלות כרוניות כמו אי ספיקת כליות, סרטן), על כך גם במאמרי: אוכלוסיית המעי הגס חיידקים טובים, דיסביוזיס ו “שומר הגנום” גן p53
- חלק ד' – על שומר החומר הגנטי🔹🐦🔥 P53 🐦🔥 והנחיות טבעיות של הגוף ל: "החלמה".
- צירי החלמה – מבוא לסדרת מאמרים על צירי החלמה והפעלתם – צירי החלמה – מבוא למחשבה חדשה על בריאות והתחדשות מאת ירון מרגולין
- 🔹 מפת מסלולי ההחלמה – 3 צירים – כללי
- 🔹המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה – מבט עומק על 3 המסלולים המרכזיים של ההחלמה.
- 🔹 חלק א' הַקְשָׁבָה – על מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין
- חלק ב' – המסלולים הביולוגיים שמשקמים את הגוף – חלק ב', על השפע וניהול תקין שלו. או איך האותות מהמזון והתנועה הופכים לצירי החלמה וריפוי.
- 🔹חלק ג' – הַפִּעְנוּחַ – על היכולת הטבעית לקדם החלמה, איך לקרוא את בדיקות המעבדה כדי לזהות את צירי החיוניות של המסלולים שמקדמים החלמה
- חלק ה' – חלבוני ההחלמה: הרשת המולקולרית של הריפוי, ההתחדשות וההומאוסטזיס מאת ירון מרגולין
- מנגנוני תיקון פנימיים בגוף: מודל אינטגרטיבי להבנת תפקוד הכליות והמערכת כולה
- חלק ו' על צירי ההחלמה ומה שמפעיל אותם – (signal architecture) מאת ירון מרגולין
- גמישות העורקים – רפואת העתיד – חלק א'
- עורקים גמישים חלק ב', הסבר מפורט על הצירים. הביולוגיה העמוקה של גמישות העורקים מאת ירון מרגולין
- קלוטו – האם קלוטו הוא מעיין הנעורים הזורם במערה מוסתרת וסודית?
- כבד שומני ❓- הכבד כמראה מערכתית: צירי מחלה, מול צירי החלמה – מבט אל רפואת העתיד
הציר המולקולרי שנוצר מכונה:
מסלול האיתות p53 (The p53 Signaling Pathway).
והוא מהווה את הלב של ה-DDR (DNA Damage Response) – תגובת התא לנזקי DNA.
להלן סרטוט זרימה מולקולרי של הציר, המציג ומסכם את הרשום למעלה, איתור השבר (DSB) מוביל להפעלת p53, הפעלת ה"ברקס" p21, וההכרעה בין החלמה וניקוי לבין מוות תאי:
🧬 נזק ל-DNA: שבר דו-גדילי (DSB)
│
▼
🚨 הפעלת אנזימי האיתות (ATM / ATR)
│
▼
[ ניתוק מ-MDM2 ] ──► p53 מיוצב ופעיל ◄── [ עיכוב הפירוק ]
│
┌──────────────────────┼──────────────────────┐
▼ ▼ ▼
[ מסלול הברקס ] [ מסלול התיקון ] [ מסלול הניקוי ]
│ │ │
הפעלת גן p21 הפעלת חלבוני הפעלת גני מוות
│ תיקון (HR/NHEJ) (Bax / Puma)
▼ │ │
חסימת חלבוני CDK ▼ ▼
│ תיקון ה-DNA אפופטוזיס (Apoptosis)
▼ והמשך מחזור התא פירוק וניקוי עצמי נקי
עצירת מחזור התא ללא דלקת ברקמה
(Cell Cycle Arrest)
מהו בעצם שבר דו-גדילי DSB ולמה הוא מסוכן כל כך?
DSB ראשי תיבות באנגלית של סוג הנזק הספציפי שנגרם ל-DNA: Double–Strand Break ובעברית: שבר דו–גדילי
🧬 מבנה ה-DNA ומשמעות השבר
מולקולת ה-DNA בנויה כסולם בעל שני גדילים השזורים זה בזה (סליל כפול).
- SSB (Single-Strand Break): אם רק גדיל אחד של הסולם נחתך, המצב פשוט יחסית. לתא יש עדיין את הגדיל השני והשלם כ"תבנית" ההנחיות לכל מה שהגוף זקוק לתפקודו, והוא יכול להעתיק ממנו את המידע המרוכז שבגדיל בקלות כדי לתקן את החלק שנשבר.
- DSB (Double-Strand Break): זהו המצב שבו שני הגדילים המקבילים נחתכים באותו מקום. במצב זה, ציר ה-DNA פשוט נקטע לשני שברים נפרדים.
⚠️ מדוע DSB הוא הנזק המסוכן ביותר לתא?
כאשר מתרחש שבר דו-גדילי DSB, נוצרות שתי בעיות קשות:
- אובדן תבנית מקומית: אין לתא גדיל מקביל וצמוד שאפשר להעתיק ממנו מיידית את המידע החסר.
- סכנת אובדן כרומוזומים: אם השברים יתרחקו זה מזה, התא עלול לאבד מקטעי גנים שלמים בזמן חלוקת התא, או לחבר בטעות קצוות של כרומוזומים שונים (מה שמוביל לטרנסלוקציות מסרטנות).
בגלל הדרמטיות של נזק מסוג DSB, התא פיתח את שתי מערכות החירום הראשיות שדיברנו עליהן: NHEJ (שפשוט מדביק את הקצוות מהר כדי שהכרומוזום לא יתפרק, גם במחיר של טעויות) ו-HR (שמחפש את הכרומטידה האחות השלמה כדי להעתיק ממנה את המידע האבוד בצורה מושלמת).
שבר דו־גדילי (DSB) הוא מצב שבו שני הגדילים של ה־DNA נחתכים.
זהו נזק מסוכן במיוחד משום שהוא עלול לגרום ל:
- אובדן מידע גנטי
- טרנסלוקציות כרומוזומליות
- מוטציות
- הזדקנות תאית
- מוות תאי או סרטנות אם התיקון כושל
לכן התא חייב להגיב מהר — אבל לא תמיד באותה דרך.
1) רקומבינציה הומולוגית (Homologous Recombination, HR)
זהו מסלול מדויק מאוד, משום שהוא משתמש בעותק תקין של אותו אזור ב־DNA כתבנית לתיקון — בדרך כלל הכרומטידה האחות.
מתי HR פועל?
בעיקר בשלבי S ו־G2 של מחזור התא, כאשר ה־DNA כבר שוכפל וקיימת כרומטידה אחות זמינה.
הרעיון הבסיסי
במקום "להדביק" את הקצוות, התא אומר:
יש לי עותק תקין של הרצף — אשתמש בו כדי לבנות מחדש את הקטע השבור בדיוק.
שלבי HR בפשטות
א. זיהוי השבר
חלבונים מזהים את ה־DSB ומגייסים את מנגנון התגובה לנזק DNA.
אחד הקומפלקסים הראשונים הוא:
- MRN complex = MRE11 + RAD50 + NBS1
קומפלקס זה:
- מזהה את השבר
- מייצב את הקצוות
- מגייס קינאזות בקרה כמו ATM
- פותח את הדרך לעיבוד הקצוות
ב. ריסוק/עיבוד הקצוות (End Resection)
כדי לבצע HR, התא צריך לייצר קצוות חד־גדיליים של 3'.
לשם כך הוא "מכרסם" את קצות ה־DNA השבור.
מי משתתף?
- MRE11
- CtIP
- EXO1
- DNA2 יחד עם BLM
התוצאה:
- נוצרים זנבות חד־גדיליים מצופי RPA
ג. החלפת RPA ב־RAD51
כעת צריך להכין את החד־גדיל ל"חיפוש" התבנית התקינה.
RPA מוחלף ב־RAD51, בעזרת מתווכים חשובים כגון:
- BRCA1
- BRCA2
- PALB2
RAD51 יוצר סיב על ה־DNA החד־גדילי ומבצע:
- חיפוש רצף הומולוגי
- פלישה לכרומטידה האחות
ד. פלישת גדיל (Strand invasion)
הגדיל השבור פולש לתוך ה־DNA התקין של הכרומטידה האחות ויוצר מבנה הקרוי D-loop.
כאן הוא משתמש ברצף התקין כתבנית.
ה. סינתזת DNA חדשה
DNA פולימראז מאריך את הקצה השבור לפי המידע התקין.
כלומר, התא משחזר את המידע שאבד במקום רק להדביק קצוות.
ו. סיום ופתרון המבנים
לאחר הסינתזה, המבנים שנוצרו מפורקים/נפתרים בכמה דרכים אפשריות:
- SDSA – מסלול נפוץ שמחזיר את הגדיל למקומו בלי crossover
- יצירת double Holliday junction
- פירוק/פתרון junctions על ידי אנזימים ייעודיים
יתרונות HR
- מדויק מאוד
- משחזר מידע שאבד
- מפחית מוטציות
- חשוב במיוחד לשמירה על יציבות הגנום
חסרונות HR
- איטי ומורכב יותר
- דורש תבנית הומולוגית
- תלוי בשלב מחזור התא
2) חיבור קצוות לא הומולוגי (Non-Homologous End Joining, NHEJ)
זהו מסלול מהיר יותר, שפועל גם כשאין כרומטידה אחות זמינה.
כאן התא לא מחפש עותק תקין; הוא מנסה להצמיד מחדש את קצות ה־DNA השבור.
מתי NHEJ פועל?
- לאורך רוב מחזור התא
- במיוחד ב־G1
- בתאים שאינם מתחלקים במהירות
- כשצריך תיקון מהיר
הרעיון הבסיסי
במקום לומר "נעתיק מן התבנית", התא אומר:
בוא נחזיק את שני הקצוות, נעבד אותם מעט אם צריך, ונדביק.
שלבי NHEJ
א. זיהוי הקצוות
החלבון המרכזי הראשון הוא:
- Ku70/Ku80
הוא נקשר לקצוות השבורים כמו טבעת/מהדק ומגן עליהם.
ב. גיוס DNA-PKcs
Ku מגייס את:
- DNA-PKcs
יחד הם יוצרים את DNA-PK complex, שמקרב את הקצוות ומארגן את התיקון.
ג. עיבוד הקצוות אם צריך
לא תמיד הקצוות "נקיים". לפעמים יש:
- בסיסים פגועים
- קצוות לא תואמים
- שיירים כימיים חוסמים
לכן התא עשוי להפעיל:
- Artemis – חיתוך/עיבוד קצוות
- פולימראזות כמו Pol μ / Pol λ
- אנזימי עיבוד נוספים
ד. ליגציה – הדבקה מחדש
בסוף מבצעים איחוי באמצעות:
- DNA ligase IV
- XRCC4
- XLF
- ולעיתים גם PAXX
יתרונות NHEJ
- מהיר
- אינו דורש תבנית
- חיוני מאוד להישרדות התא
- חשוב בתאים שאינם בשלב שכפול
חסרונות NHEJ
- עלול להיות לא מדויק
- לעיתים מאבד/מוסיף כמה נוקלאוטידים
- עלול ליצור indels
- אם הקצוות הלא נכונים מתחברים זה לזה — ייתכנו שינויים כרומוזומליים
ההבדל העקרוני ביניהם במשפט אחד
- HR = “נשחזר את המקור לפי עותק מדויק.”
- NHEJ = “נסגור מהר את הקרע, גם אם התפר לא מושלם.”
השוואה בין שני המסלולים
| מאפיין | HR | NHEJ |
|---|---|---|
| דיוק | גבוה מאוד | בינוני–נמוך יחסית |
| צורך בתבנית | כן, כרומטידה אחות | לא |
| שלב מחזור התא | בעיקר S/G2 | בעיקר G1, אך לא רק |
| מהירות | איטי יותר | מהיר יותר |
| סיכון למוטציות | נמוך | גבוה יותר |
| שימוש עיקרי | תיקון נאמן | תיקון הישרדותי מהיר |
איך התא בוחר בין HR ל־NHEJ?
זו שאלה מרכזית מאוד. הבחירה אינה מקרית.
גורמים עיקריים:
1. שלב מחזור התא
- G1 → לרוב NHEJ
- S/G2 → עולה הסיכוי ל־HR בגלל קיום כרומטידה אחות
2. מידת עיבוד הקצוות
ברגע שמתחיל end resection משמעותי, התא כבר "נוטה" לכיוון HR.
לכן הרגע שבו התא מחליט אם לכרסם את הקצוות הוא נקודת הכרעה חשובה.
3. חלבוני בקרה
- 53BP1 נוטה לעודד NHEJ על ידי הגבלת resection
- BRCA1 מקדם resection ולכן דוחף לכיוון HR
כלומר, יש כאן מעין מאבק רגולטורי:
- 53BP1 = לשמור קצוות, לסגור מהר
- BRCA1 = לפתוח ולעבד, כדי לתקן במדויק
ומהו alt-EJ / MMEJ?
מעבר לשני המסלולים הקלאסיים, יש גם מסלולי גיבוי, ובעיקר:
Microhomology-Mediated End Joining (MMEJ)
זהו מסלול ביניים/גיבוי שבו התא משתמש במקטעי הומולוגיה קצרים מאוד (microhomology), לרוב של כמה נוקלאוטידים, כדי לחבר את הקצוות.
מאפיינים:
- דורש עיבוד קצוות
- פחות מדויק מ־HR
- לרוב יוצר מחיקות
- קשור לעיתים ל־POLQ (DNA polymerase theta)
המסלול הזה חשוב במיוחד כש־HR או NHEJ הקלאסי פגועים.
חלבוני מפתח שכדאי להכיר
ב־HR
- MRN (MRE11-RAD50-NBS1) – זיהוי ותחילת תגובה
- ATM – אות נזק ובקרת תיקון
- CtIP – התחלת resection
- EXO1 / DNA2 / BLM – הרחבת resection
- RPA – קושר ssDNA
- BRCA1 / BRCA2 / PALB2 – הכוונת HR
- RAD51 – לב המסלול; חיפוש הומולוגיה ופלישת גדיל
ב־NHEJ
- Ku70/Ku80
- DNA-PKcs
- Artemis
- XRCC4
- Ligase IV
- XLF / PAXX

כיצד נזק גנומי מתמשך בכליה דוחף את התא מהתחדשות לשיתוק, לפיברוזיס ולאובדן תפקוד
במחלת כליות כרונית, ובמידה מסוימת גם לאחר פגיעת כליה חריפה, מצטבר לעיתים עומס של נזקי DNA. כשהנזק אינו מתוקן היטב — התא לא רק "מסתכן בסרטן", אלא גם עלול להיכנס להזדקנות תאית, לעצור חלוקה, למות, להפריש אותות דלקתיים, ולדחוף את הכליה לכיוון של פיברוזיס ואובדן נפרונים [מקור]. כלומר, בכליה הנזק הגנומי מתבטא לא רק כממאירות, אלא גם כהחמרה מבנית ותפקודית של המחלה עצמה [מקור1, מקור2, מקור3].
הבנת הגורמים והקשרים בין תרופות, תזונה שגויה (יתר חלבונים, סוכר ופחמימות), לחץ נפשי לבין נזק ל-DNA הם קריטים להבנת המזון כתרופה, הפעלת צירי החלמה הם חזית המחקר כיום.
שברים דו-גדיליים (DSBs) נגרמים משני מקורות עיקריים: נזק סביבתי (חיצוני) ותהליכים פנימיים של הגוף. להלן הפירוט המדעי המדויק לגבי הגורמים שנחקרים כיום כמי שגורמים ומובילים לנזק שמקדם חולי:
🥩 1. תזונה ו-TMAO (מטבוליזם של בשר אדום)
כאשר אנו אוכלים בשר אדום, ביצים או מוצרי חלב שמנים, חיידקי המעיים מפרקים רכיבים כמו כולין וקרניטין לחומר בשם TMA. חומר זה מגיע לכבד והופך ל-TMAO (Trimethylamine N-oxide).
- המנגנון ל-DSB: רמות גבוהות של TMAO בדם מעוררות עקה חמצונית (Oxidative Stress) קשה בתאים, במיוחד בתאי דופן כלי הדם (אנדותל).
- העקה החמצונית מייצרת רדיקלים חופשיים (ROS) בעודף עצום. רדיקלים אלו תוקפים ישירות את קשרי הפוספט-סוכר ב-DNA, ומסוגלים לגרום לחיתוך סימולטני של שני הגדילים – כלומר, יצירת DSB.
🧬 2. יתר הומוציסטאין (Hyperhomocysteinemia)
🔬 המנגנון המולקולרי לפגיעה ב-DNA:
הציבור מפספס בדרך כלל את הסכנות שבהומוציסטאין גבוה.
הומוציסטאין ברמות גבוהות בדם פועל כרעל תאי חריף בכמה מישורים:
- עקה חמצונית ישירה: הומוציסטאין עובר תהליך של חמצון עצמי (Auto-oxidation) בדם. תהליך זה מייצר כמויות אדירות של רדיקלים חופשיים (ROS) מסוג מימן על-חמצני (Hydrogen Peroxide). רדיקלים אלו תוקפים ישירות את קשרי הפוספט של ה-DNA וגורמים ל-DSB.
- חסימת מנגנוני התיקון: הומוציאסטין משבש את תהליך המתילציה בגוף. ללא מתילציה תקינה, התא אינו מצליח לייצר או להפעיל חלבוני מפתח האחראיים על בקרת איכות ותיקון ה-DNA (כמו p53 או אנזימי תיקון השברים. ה-p53 עצמו מופעל בעיקר על ידי זרחון (Phosphorylation) ואיצטילציה.), מה שמשאיר את הגנום פגיע וחשוף.
🥩 המקורות שמקדמים את הסינתזה ההרסנית של הומוציסטאין:
הומוציסטאין אינו מגיע ישירות מהמזון, אלא מסונתז בגוף מחומצת האמינו מתיונין (Methionine). המקורות המרכזיים שמקפיצים את רמותיו הם:
- תזונה עשירה מדי במתיונין (חלבון מן החי):
- בשר אדום ובשר בקר שמן (המקור המרוכז ביותר).
- עוף והודו (בצריכה מוגברת).
- ביצים ומוצרי חלב קשים/שמנים.
- דגים מסוימים (כמו טונה וסלמון).
- ה"שותפים לפשע" – מחסור בוויטמינים (מניעת פירוק):
הסינתזה הופכת להרסנית ומצטברת בעיקר כאשר חסרים בגוף ה"מנקים" של ההומוציאסטין. מחסור ברכיבים הבאים מונע מהגוף לנטרל אותו:- מחסור בוויטמין B12 ובוויטמין B6 (נפוץ אצל טבעונים, צמחונים או אנשים עם בעיות ספיגה במערכת העיכול).
- מחסור בחומצה פולית (B9) (נפוץ עקב תזונה דלה בעלים ירוקים, או עקב פגם גנטי נפוץ באנזים MTHFR המונע את ניצול הוויטמין).
- שילובים תזונתיים מורכבים (כמו חומוס ופחמימה):
חלבונים מן הצומח (כמו קטניות/חומוס) הם אמנם דלים במתיונין בפני עצמם, אך כאשר משלבים אותם עם דגנים (כמו פיתה, לחם או אורז), נוצר חלבון מלא. השילוב הזה מעלה באופן מיידי את זמינות המתיונין בגוף, ואצל אנשים עם רגישות גנטית או מחסור בוויטמיני B, הוא עלול לתרום להאצת מסלול ההומוציסטאין.
💼 3. לחץ נפשי (קריירה דורשנית) ויתר קורטיזול
הקשר בין סטרס פסיכולוגי כרוני לבין פגיעה פיזית ב-DNA הוא מוכח מחקרית. כאשר אדם נמצא בלחץ מתמיד, בלוטת יותרת הכליה מפרישה רמות גבוהות של קורטיזול וקולטנים מופעלים באופן כרוני.
- המנגנון ל-DSB: קורטיזול גבוה לאורך זמן משבש את ביטויים של גנים האחראיים על בקרת איכות ותיקון ה-DNA.
- בנוסף, סטרס כרוני מקצר את הטלומרים (קצוות הכרומוזומים). כאשר הטלומרים הופכים קצרים מדי, קצוות הכרומוזום נחשפים, והתא מזהה את הקצה הפתוח בטעות כ-DSB. זה מפעיל אזעקת שווא תאית שמובילה להזדקנות התא (Senescence) או למותו.
💊 4. תרופות וטיפולים רפואיים
חלק מהתרופות מייצרות שבר דו-גדילי DSB בכוונה תחילה, וחלק כתוצאת לוואי:
- תרופות כימותרפיות: תרופות כמו דוקסורוביצין או אטופוסיד הן מעכבי האנזים טופואיזומראז (Topoisomerase). תפקידו הטבעי של האנזים הוא לחתוך את ה-DNA זמנית כדי לשחרר מתח בזמן שכפול, ולחבר בחזרה. התרופות "תוקעות" את האנזים כשה-DNA חתוך, ובכך יוצרות DSB מכוון כדי להרוג את תאי הסרטן.
- אנטיביוטיקות מסוימות: קבוצת הציפרופלוקסצינים (fluoroquinolones) פועלת במנגנון דומה על חיידקים, אך במינונים גבוהים או ברגישות גבוהה עלולה להגביר עקה חמצונית ותתי-נזקים ב-DNA של המיטוכונדריה בתאי אדם.
☀️ גורמים קלאסיים נוספים שחשוב להכיר:
- קרינה מייננת (קרינת רנטגן, קרינה קוסמית): פוגעת ישירות בליבת הגנום וקורעת את שני הגדילים במכה אחת.
- קרינת UV (שמש): מייצרת בעיקר נזקים חד-גדיליים (SSB), אך אם התא מנסה לשכפל DNA פגום כזה בשלב ה-S, מזלג השכפול קורס והופך מיד ל-DSB.
למה זה חשוב כל כך ברפואה ובביולוגיה של סרטן?
מפני שכאשר מסלולי תיקון DNA נפגעים, התא מתחיל לאבד שליטה על הגנום.
דוגמאות:
BRCA1 / BRCA2
פגיעה בגנים אלה מחלישה HR, ולכן התאים נאלצים להישען יותר על מסלולים פחות מדויקים.
זה מעלה חוסר יציבות גנומית ומגביר סיכון לסרטן.
PARP inhibitors
בתאים עם חסר ב־HR (למשל פגיעה ב־BRCA), אפשר לנצל זאת טיפולית באמצעות מעכבי PARP.
זו דוגמה יפה ל־synthetic lethality:
- התא הסרטני כבר חלש בתיקון DNA
- חוסמים לו מסלול תיקון נוסף
- התא קורס
דרך אינטואיטיבית לזכור
דמיין ספר עתיק שנקרע באמצע:
HR
יש לך עותק זהה של הספר לידך.
אתה משחזר כל מילה בדיוק לפי העותק.
NHEJ
אין עותק.
אתה מצמיד את שני חצאי הדף, מדביק, ואולי גוזר מעט שוליים כדי שיתאימו.
ואם נרצה לדייק עוד יותר: HR הוא לא מסלול יחיד
תחת הכותרת "HR" יש כמה תתי־מסלולים:
- DSBR (Double-strand break repair model)
- SDSA (Synthesis-dependent strand annealing)
- BIR (Break-induced replication)
- תיקון של stalled forks בשכפול
כלומר, HR הוא משפחה של מנגנונים המשתמשים בהומולוגיה כדי לשחזר מידע.
מסקנה קצרה
כאשר ה־DNA נשבר בשני הגדילים, התא עומד בפני בחירה בין שני עקרונות יסוד:
- תיקון מדויק באמצעות תבנית → HR
- איחוי מהיר של הקצוות → NHEJ
הבחירה תלויה ב:
- שלב מחזור התא
- זמינות כרומטידה אחות
- מצב הקצוות
- חלבוני בקרה כמו BRCA1 ו־53BP1
חלק ב'

רפואת העתיד – תהליכי החלמה ללא תרופות וחומרים סינטטיים
אין כיום דרך פשוטה "להדליק" באופן ממוקד את HR או את NHEJ כרצוננו, אך בהחלט ישנם צירים לא־תרופתיים שעשויים להפחית את עומס הנזק ל־DNA ולתמוך בסביבה התאית הנחוצה לתיקון תקין: הפחתת עקה חמצונית, שיפור תפקוד מיטוכונדריאלי, שימור מאגרי NAD⁺, תמיכה בפעילות סירטואינים כגון SIRT1 ו־SIRT6, שמירה על יציבות אפיגנטית, והגנה על טלומרים. במילים אחרות, במקום "להכריח" את התא לתקן, אפשר לנסות לשפר את התנאים שבתוכם מנגנוני התיקון פועלים.
— מבט עומק בתהליכי החלמה
כיצד יוצרים בתא תנאים שבהם התא בוחר תיקון, התחדשות והישרדות — ולא סנסנס (תאי "זומבי"), פיברוזיס וקריסה
שבר דו־גדילי ב־DNA איננו רק פגיעה מבנית במולקולת התורשה. זהו רגע של הכרעה ביולוגית. מרגע שנפער השבר, התא נדרש לא רק להפעיל מנגנון תיקון מתאים, אלא גם להכריע אם בידו עדיין הכוח, המשאבים והיציבות הפנימית הנחוצים לשיבה אל מסלול של חיים. לעיתים התשובה היא תיקון, התאוששות והמשך תפקוד; אך כאשר הנזק מצטבר על רקע עקה חמצונית, דלקת כרונית, דלדול אנרגטי, שחיקה אפיגנטית ופגיעה מיטוכונדריאלית — השבר הגנומי חדל להיות אירוע נקודתי והופך לנקודת מפנה, שממנה התא עלול לגלוש אל עצירת מחזור, הזדקנות תאית, הפרשת אותות דלקתיים ופיברוזיס.
במובן זה, שאלת התיקון הגנומי רחבה יותר מן השאלה כיצד מחברים מחדש שני קצוות שבורים של DNA.
כדי שתא יבחר בהחלמה, עליו להיות מצוי במצב ביולוגי המאפשר תיקון:
עליו לשמר כרומטין בר־ארגון, אספקת אנרגיה מספקת, הגנה מפני שטף מתמשך של רדיקלים חופשיים, מאגרי NAD⁺ שאינם קורסים, ומערכות בקרה המסוגלות להבחין בין נזק חולף לבין מצב שאין ממנו חזרה. ללא תנאים אלה, גם מסלולי תיקון משוכללים עלולים לפעול באיטיות, בחוסר דיוק, או להידחק מפני תוכניות תאיות אחרות — עצירת חלוקה, סנסנס (או תאי "זומבי"), אפופטוזיס, ולעיתים גם הישרדות פגומה של תא שנפגע [מקור1, מקור2, מקור3, מקור4, מקור5].
הערה: NAD⁺ (ניקוטינאמיד אדנין דינוקלאוטיד) הוא קואנזים חיוני הנמצא בכל תא בגוף האדם. הוא מתפקד כ"נשא אלקטרונים" ומשחק תפקיד ליבה בייצור אנרגיה במיטוכונדריה (הפיכת מזון ל-ATP), בתיקון נזקי DNA, ובוויסות תהליכי הזדקנות ובריאות התא. בלעדי NAD⁺ (, תהליכי חילוף החומרים הבסיסיים המפיקים אנרגיה בגוף לא היו מתקיימים.
הדבר נכון במיוחד בכליה. זוהי רקמה בעלת דרישה מטבולית גבוהה, חשיפה ממושכת לעקה חמצונית, לרעלנים אורמיים, לאיסכמיה, לדלקת ולשיבושי מיטוכונדריה. בתנאים אלה, נזקי DNA אינם רק "סמן" לפגיעה, אלא מנגנון פעיל הדוחף את הרקמה לעבר אובדן נפרונים, עצירת התחדשות, מעבר של תאי אפיתל ומזנכימה למצב מפריש־דלקת, והצטלקות מתקדמת של הפרנכימה. לכן,
במחלת כליות כרונית, הדיון ב־DSB אינו יכול להסתיים במסלולי HR ו־NHEJ בלבד;
הוא חייב לעבור אל השאלה:
מה נדרש כדי שתא כלייתי יישאר תא בר־תיקון, ולא יהפוך לתא מזדקן שמזין את תהליך הפיברוזיס.
מכאן עולה עיקרון חשוב: אין בידינו כיום דרך פשוטה "לכפות" על התא לבחור במסלול תיקון אידיאלי, אך ישנם צירים ביולוגיים, מסלולי החלמה ותיקון שעשויים להשפיע עמוקות על סביבת ההחלטה שבתוכה התיקון מתרחש. צירים אלה נחקרים כיום, ככל שהמחקר מעמיק גדלה הבנתנו שהפעלתם של צירי ההחלמה נקשרת בשיקום, ניקוי רעלים וברפואת העתיד. צירי ההחלמה, חלבוני השיקום נועדו על ידי כוחות הטבע למטרות של ריפוי, אך הם גם עשויים רק להפחית את עומס הנזק, לייצב את הכרומטין, לשמר אנרגיה תאית, לעכב הזדקנות מוקדמת, ולהגדיל את הסיכוי שהתגובה לנזק תסתיים בתיקון ובהישרדות ולא בקריסה. במילים אחרות, אם חלקו הראשון של המאמר עסק בשאלה כיצד התא מתקן שבר, חלק זה של המאמר עוסק בתנאים המוקדמים המאפשרים לתא להיות בכלל מסוגל לתקן.
הציר הראשון והעמוק ביותר בדיון זה הוא הטלומרים. קצות הכרומוזומים מציבים בפני התא את אחת הבעיות היסודיות ביותר של תחזוקת הגנום: כיצד להגן על מבנה הדומה לשבר דו־גדילי, מבלי להפעיל עליו בטעות את מנגנוני התיקון של שבר אמיתי.
כאשר ההגנה הטלומרית נשחקת, הגבול בין קצה כרומוזום תקין לבין אות מצוקה גנומי מיטשטש, והתוצאה עלולה להיות הפעלה כרונית של תגובת נזק ל־DNA, סנסנס, אי־יציבות כרומוזומלית ופגיעה ביכולת ההתחדשות של הרקמה.
משום כך, הדיון בתנאים המאפשרים לתא לבחור בתיקון ובהחלמה חייב להתחיל בטלומרים — בגבול הדק שבין הגנה על החיים לבין תחילת ההזדקנות התאית.
הערה: תאי סנסנס (Senescent cells) הם תאים מזדקנים שאינם מתחלקים עוד אך מסרבים למות. תאים אלו גורמים ל"נזק ביולוגי" מצטבר בכך שהם מפרישים חומרים רעילים המעוררים דלקת כרונית, פוגעים בתאים בריאים סמוכים, ומאיצים תהליכי זדקנות ומחלות [מקור1, מקור2, מקור3, מקור4].
חלק ג – המשך בקישור למטה.
חלק ג'
על הטלומרים:
- מדוע קצה כרומוזום דומה ל־DSB
- Shelterin וההגנה מפני זיהוי שגוי של הטלומר כשבר
- שחיקת טלומרים: DDR, סנסנס, איחויי קצה־לקצה ואי־יציבות גנומית
- טלומרים בכליה: הזדקנות תאית, אובדן יכולת התחדשות ופיברוזיס
- אסטרגלוס ונגזרותיו (למשל cycloastragenol / TA-65) הפעלת טלומראז — הארכת טלומרים ומניעת שבר דו-גדילי עקב קיצור טלומרים: ההבטחה והסיכון
המשך בחלק ג'
נשארו לך שאלות
🔬אשמח להשיב על כל שאלה
בבקשה לא להתקשר משום שזה פשוט לא מאפשר לי לעבוד – אנא השתמשו באמצעים שלפניכם
למען הסר ספק, חובת התייעצות עם רופא (המכיר לפרטים את מצבו הבריאותי הכללי של כל מטופל או שלך) לפני שימוש בכל תכשיר, מאכל, תמצית או ביצוע כל תרגיל. ירון מרגולין הוא רקדן ומבית המחול שלו בירושלים פרצה התורה כאשר נחשפה שיטת המחול שלו כבעלת יכולת מדהימה, באמצע שנות ה – 80 לרפא סרטן. המידע באתר של ירון מרגולין או באתר "לחיצות ההחלמה" (בפיסבוק או MARGOLINMETHOD.COM ), במאמר הנ"ל ובמאמרים של ירון מרגולין הם חומר למחשבה – פילוסופיה לא המלצה ולא הנחייה לציבור להשתמש או לחדול מלהשתמש בתרופות – אין במידע באתר זה או בכל אחד מהמאמרים תחליף להיוועצות עם מומחה מוכר המכיר לפרטים את מצבו הבריאותי הכללי שלך ושל משפחתך. מומלץ תמיד להתייעץ עם רופא מוסמך או רוקח בכל הנוגע בכאב, הרגשה רעה או למטרות ואופן השימוש, במזונות, משחות, תמציות ואפילו בתרגילים, או בתכשירים אחרים שנזכרים כאן.
physician (who knows in detail the general health of each patient or yours) before using any medicine, food, extract or any exercise. The information on Yaron Margolin's website or the "Healing Presses" website (on Facebook or YARONMARGOLIN.COM), in the above article and in Yaron Margolin's articles are material for thought – philosophy neither recommendation nor public guidance to use or cease to use drugs – no information on this site or anyone You should always consult with a qualified physician or pharmacist regarding pain, bad feeling, or goals and how to use foods, ointments, extracts and even exercises, or other remedies that are mentioned as such
מאמרים אחרונים
- שבר דו־גדילי ב־DNA: מסלולי התיקון, בחירת התא בין HR ל־NHEJ, והשלכות על סרטן, הזדקנות ומחלת הכליות, חלק א' וחלק ב'/ ירון מרגולין
- שבר דו־גדילי ב־DNA: מסלולי התיקון, בחירת התא בין HR ל־NHEJ, והשלכות על סרטן, הזדקנות ומחלת הכליות / חלק ג' על ההחלמה ומה ניתן לעשות❓
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ב', על הפוטנציאל הקיים בגוף האדם לקדם תהליכי החלמה מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ג', על התקדמותם של צירי ההחלמה בגוף האדם
- 🔷 המסלולים הביולוגיים שמשקמים את הגוף: p53 — שומר הגנום – חלק ד’ — ירון מרגולין
- חלק ה' – חלבוני ההחלמה: הרשת המולקולרית של הריפוי, ההתחדשות וההומאוסטזיס מאת ירון מרגולין
- חלק ו' על צירי ההחלמה ומה שמפעיל אותם – (signal architecture) מאת ירון מרגולין
- מנגנוני תיקון פנימיים בגוף: מודל אינטגרטיבי להבנת תפקוד הכליות והמערכת כולה – 3 צירי ההחלמה המרכזיים
- המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה
- גמישות העורקים. חלק א' על צירי המחלה מול צירי ההחלמה – הסיפור האמיתי של כלי הדם
- חלק ב' גמישות העורקים, מה אפשר לעשות
- ⭐ התחדשות כלייתית: מקרה נדיר של ריפוי טבעי — ומה שהוא מלמד את כולנו
- דרכים חדשות לוויסות יעיל של טריגליצרידים (Triacylglycerols) בדם
- אי ספיקת כליות – טיפול ללא תרופות
- על ההחלמה – למה הגוף לא מחלים — וכיצד להחזיר לו את היכולת הטבעית לעשות זאת
- שינה חשובה להחלמה – 🌜זמן הריפוי החשוב מתרחש בלילה
- מיומטי תנועה – התקווה החדשה לחולי כליות: תרכובות שמחקות את צירי ההחלמה של פעילות גופנית
- על הגאונות והגאון
- 📘 The Biological Pathways That Restore the Body: A New Perspective on Recovery
- הגוף יודע על המחלה: כיצד לזהות אותה באמצעות מבט עומק בבדיקות המעבדה ובתסמינים הגופניים
- ברומלין – קמח קליפות אננס, חומץ אננס – רוצה לדעת כיצד להכין תרופה ביתית, אנטי-דלקתי מקליפות האננס? – .ירון מרגולין – מייסד לחיצות ההחלמה להבראת הגוף והנפש
(opens in a new tab) - דיאליזה: מאבק הירואי שאינו ריפוי
- רעלים אורמיים מקדמים דיאליזה – הוכח לאחרונה שרעלנים אורמיים קשורים למיקרוביוטה של המעי הגס – איך להפעיל מנגנוני שיקום טבעיים בגוף ללא תרופות ולהימנע מדיאליזה.
- מכתב תודה ממחלימת כליות בתוך ארבעה חודשים.
- למה חולים שנוטלים כדורים להפחתת לחץ דם גבוה, נדרשים להפעיל מנגנוני שיקום טבעיים בגוף כדי לעקב דיאליזה
- איך לא להגיע לדיאליזה
- תובנות חדשות אין להתחיל דיאליזה לפני בדיקת – מצבו של ציר FGF23-Klotho
- קלוטו – האם קלוטו הוא מעיין הנעורים הזורם במערה מוסתרת וסודית?
- איך התפתחו לחיצות ההחלמה בבית מחול ירון מרגולין בירושלים
- המזון הבסיסי
- המזון הבסיסי והחומצי – מה כן ומה לא נכון. מה עובד על ההחלמה ומה הן אגדות? חומציות מגיעה מתזונה והיא קמה והורסת את הכליות
- תובנות חדשות אין להתחיל דיאליזה לפני בדיקת – מצבו של ציר FGF23-Klotho
- עיגול פינות מקשה על החלמת הכליות. מיומנו של מאסטר בהחלמת הכליות – הכליות לא סולחות על הזנחה:
- תובנות חדשות אי-ספיקת כליות תלויה במצבו של ציר FGF23-Klotho
- דיאליזה: מאבק הירואי שאינו ריפוי
- פחות מתים – טכניקת דיאליזה חדשה שבולמת את קצב המתים ומגבירה את ההישרדות בכשל כליות.
- שכיחות של דיכאון וחרדה בקרב חולי כליות.
- המזון הבסיסי והחומצי – מה כן ומה לא נכון. מה עובד על ההחלמה ומה הן אגדות? חומציות מגיעה מתזונה והיא קמה והורסת את הכליות
- הביוכימיה של ההחלמה: מפת העל של החיים
- שבריריות גופנית מחדל
- המיקרוביום הכלייתי (האורוביום Urobiome) נחשף
- ניקוי כבד טבעי – הדרך הפשוטה לבריאות מתחדשת
- מיומטי תנועה – התקווה החדשה לחולי כליות: תרכובות שמחקות את צירי ההחלמה של פעילות גופנית
- הגוף יודע על המחלה: כיצד לזהות אותה באמצעות מבט עומק בבדיקות המעבדה ובתסמינים הגופניים
- איך התפתחו לחיצות ההחלמה בבית מחול ירון מרגולין בירושלים
- דרכים חדשות לוויסות יעיל של טריגליצרידים (Triacylglycerols) בדם
- מלטונין הוכח שהורמון השינה נוגד חמצון רב עוצמה ויעיל כתרופה בקרב חולי כליה
- מיטוכונדריה – במיטוכונדריון מתחילה מחלת הכליות
- הביוכימיה של ההחלמה: מפת העל של החיים
- אבנים בכליות – אוקסלטים, היפרוקסלוריה ויתר חומצה אוקסלית – רוצה להפעיל מנגנוני שיקום טבעיים בגוף האדם ללא תרופות?
- זרימת דם כלייתית
- השפעות של הורמון הגדילה (GH) על תפקוד הכליות בבריאות
- השמנה בטנית – סטרס ביולוגי — מנגנון העל שמסביר השמנה בטנית גם בלי אכילת יתר. למה לא יורדים בבטן בתהליכי הרזיה מסודרים?
- גלוטתיון – מנוע התיקון המרכזי של הגוף
- למה חולים שנוטלים כדורים להפחתת לחץ דם גבוה, ידרשו להפעיל צירי החלמה ביולוגיים אם הם רוצים להתגבר על המסלול הרץ אל תוך הדיאליזה
- איך לא להגיע לדיאליזה
- תובנות חדשות אין להתחיל דיאליזה לפני בדיקת – מצבו של ציר FGF23-Klotho
- תובנות חדשות אי-ספיקת כליות תלויה במצבו של ציר FGF23-Klotho
- פחות מתים – טכניקת דיאליזה חדשה שבולמת את קצב המתים ומגבירה את ההישרדות בכשל כליות.
- שכיחות של דיכאון וחרדה בקרב חולי כליות
- הערכות שונות במדידת אשלגן בפלזמה שוללות לפעמים שלא בצדק יתר אשלגן בה – היפרקלמיה פסאודו היפרקלמיה – Hyperkalemia
- מדד חדש לבריאותך – ביטול האסרוסקלרוזים
- אי ספיקת כליות – טיפול ללא תרופות
- אוראה – למה האוראה מוגברת ומופחת בדם Urea
- Kidney Failure Taming the Rogue Kidneys Vital Information
- על הגאונות והגאון
- שיחות ההחלמה ושיחות בין ידידים – על ההבדלים ביניהן
- הפסיכולוגיה הטיפולית וההוליסטית – תולדות הפסיכותרפיה
- תרופות הרגעה שמשאירות אותך רעב וחרד – ציפּרָלֵקס, פּרוֹזַק, פלואוקסטין
- מגילת רות האלמנה שיצאה ממרחב הנוחות
- יש פתרון כולל לבעיות הקשורות לדימוי עצמי, חוסר בטחון וערך עצמי נמוך – דופמין
- חרדה
- איך מורידים לחץ וחרדות באופן טבעי? התמודדות עם סטרס ולחץ נפשי
- טיפול נפשי ללא תרופות – חרדות, פחדים וייאוש – תסמינים שיש להם פתרון טבעי, מדויק ומהיר
- למה אני לא מצליח להתמיד – והסוכר
- גמישות היא מצב נפשי – אתגר בזרימה ושינוי – אני מבקש להתגמש
- חזרה לבסיס – עקרונות צירופי המזון
- אי ספיקת כליות
- PGC-1α, יעד טיפולי חדש נגד מחלת הכליות
- התערבויות בניהול רגשות, מחשבות ואורח חיים כדי להתמודד עם מגיפה של מחלות כליתיות
- פורינים חוץ תאיים ותחזוקת הכליות
- רככת היפופוספטמית (זרחן נמוך מהנורמה) – מחלת כליות כרונית – או הפרעת עצם מינרלית
- סלניום Se התגלה כאנטי- אייג'ינג ומגן מפני מחלות כרוניות
- מלטונין הוכח שהורמון השינה נוגד חמצון רב עוצמה ויעיל כתרופה בקרב חולי כליה
- רעלים אורמיים מקדמים דיאליזה – הוכח לאחרונה שרעלנים אורמיים קשורים למיקרוביוטה של המעי הגס – איך להפעיל מנגנוני שיקום טבעיים בגוף האדם ללא תרופות ולהימנע מדיאליזה
- מאמר הצלבת איברים – הדרך להבנת המחלה– כאן.
- מיטוכונדריה – במיטוכונדריון מתחילה מחלת הכליות
- תובנות חדשות אי-ספיקת כליות תלויה במצבו של ציר FGF23-Klotho
- מחלת שינוי מינימלי Minimal Changes Disease מינימל צ’יינג’
- מחלת כליות פוליציסטית אוטוזומלית דומיננטית (ADPKD)
- התכנית לשיקום הכליות – כאן.
- מכתב תודה ממחלימת כליות בתוך ארבעה חודשים.
- הקשר של כבד שומני, ומחלת כליות כרונית – מחקר חדש.
- מחלת כליות – תמיכה בתהליכי התאוששות – לשם כך יש לזהות אותה מוקדם ככל האפשר – הקסטסרופה!
- זיהום סביבתי מקדם את מחלת הכליות
- מחלת כליות פוליציסטית אוטוזומלית דומיננטית (ADPKD)
- השפעות של הורמון הגדילה (GH) על תפקוד הכליות בבריאות ובמחלות כליות
- כלית העל מספרית Accessory kidney
- דנרבציה כלייתית Renal denervation (RDN)
- השפעות של הורמון הגדילה (GH) על תפקוד הכליות בבריאות ובמחלות כליות
- אנמיה למה חסר לי ברזל?
- פגיעה במערכת העיכול ללא תרופות?
- קל לשמור על הכבד
- הכבד כמראה מערכתית: צירי מחלה מול צירי החלמה
- חלק ו' על צירי ההחלמה ומה שמפעיל אותם – (signal architecture) מאת ירון מרגולין
- 🔹 מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים🔹 את הגוף – חלק ב', על ההחלמה מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ג', על ההחלמה
- 🔷 המסלולים הביולוגיים שמשקמים את הגוף: p53 — שומר הגנום – חלק ד’ — ירון מרגולין
- חלק ה' – חלבוני ההחלמה: הרשת המולקולרית של הריפוי, ההתחדשות וההומאוסטזיס מאת ירון מרגולין
- המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה
- 📘 The Biological Pathways That Restore the Body: A New Perspective on Recovery
- גמישות העורקים – רפואת העתיד – חלק א'
- עורקים גמישים חלק ב', הסבר מפורט על הצירים. הביולוגיה העמוקה של גמישות העורקים מאת ירון מרגולין
- קלוטו – האם קלוטו הוא מעיין הנעורים הזורם במערה מוסתרת וסודית?
- המחמצן הגדול – Ros ודרכי ההתגוננות מפניו ללא תרופות
- חימצון האינסולין והגברה של חומצת שתן בדם, גאוט, אי ספיקת כליות, שבץ לב וסוכרת
- מיטוכונדריה – במיטוכונדריון מתחילה מחלת הכליות
- ירידה בתפקוד הכליות – מה לעשות?
- דיוקסינים במזון
- כשהמוסר מתנתק מן החיים: כיצד הפכה האידאולוגיה הנאורה מביקורת מוסרית לעמדה הרסנית ומשתקת
- קדרה ואבדון
- Cauldron and Ruin
- אוכלוסיית המעי הגס חיידקים טובים, דיסביוזיס ו “שומר הגנום” גן p53
- מחסור בברזל
- מיומטי תנועה – התקווה החדשה לחולי כליות: תרכובות שמחקות את צירי ההחלמה של פעילות גופנית
- בשר, כליות וזרימת דם מואצת: כיצד תזונה מהחי פוגעת במערכת הסינון של הגוף
- אמנות ההחלמה האבודה
- קרקר מדהים מכוסמת ירוקה – מתכון
- השפעה קלינית של ריבס על הטיפול באי ספיקת כליות כרונית: כפי שמשתקף במחקרים.
- פרנצ'סקו מסינה Francesco Messina אלהים באדם שחיפש אחר היופי
- התרופות הטובות והרע – חיסול האוצר המגן על משק המיים ולחץ הדם בגוף – האשלגן
- התרופות הטובות והרע – אבץ
- תרופות הרגעה שמשאירות אותך רעב וחרד – ציפּרָלֵקס, פּרוֹזַק, פלואוקסטין
- התרופות והרע – ביוטין B7 ויטמין H
- יתר לחץ דם
- התרופות והרע – ומי הורס לך את ה-Q10
- מחסור בויטמין בי-1 מייצר סיוטי לילה וחלומות זוועה B1
- גלוטתיון Glutathione ו-N-אצטיל-ציסטאין (NAC)
- שלושת המיצים
- על ההזנחה –
- ניתוק רגשי – על הקורוציונה
- די לכאב
- ראיית המעמקים – כניסה לטרקלין או על החיים האמתיים.
- התודעה השלילית
- ביקורת מבזה
- איך לצאת ממצבי תקיעות בחיים – שיטת שלוש השאלות בגובה העיניים
- נמאס לי מהחיים מה עושים
- על היכולת להשתקם, לקום מאבק הדרך ומכאב הפרידה
- ריגוש רעיוני או התקווה והדופמין – יחשפו בפניך כל הסודות שבהתמכרות שבהמתנה – ריגוש, המתנה, תקוה ודופמין – המאמר כאן.
- מסלול ההתפתחות הנורמטיבי בחיי אדם ודמימותיו בטבלה – סדרת מאמרים.
- שפינוזה כממשיך דרכו של אפלטון – תורת הנפש
- תורת הנפש של אפלטון – הידיעה
- האני הגמיש-לנפתלי עירוני
- לחלק ראשון של –ימי הסנאל שלי –ימי הסנאל שלי – להתגבר על אוטיזם.
- תככים ומזימות – תהליך חברתי סביב גרוש יהודי ספרד – 1
- תככים ומזימות – תהליך חברתי סביב גרוש יהודי ספרד – 2
- תככים ומזימות – תהליך חברתי סביב גרוש יהודי ספרד – 3
- הרופאים הגדולים בהיסטוריה
- אלקמאון מקרוטון (Alcmaeon of Croton) – חוקר המוח ומערכת העצבים הראשון בהיסטוריה
- פרנצ'סקו מסינה Francesco Messina אלהים באדם שחיפש אחר היופי
- פיטר דה הוך – גאון ומוביל דרך
- השבועה ההיפוקרטית – העדויות על קשרים פיננסיים של רופאים עם תעשיית התרופות, הולכות ורבות, האם השבועה מופרת כיום?
- פייר בושאן – האיש שהציל את חגיגת הנישואין של לואי ה-14 מאת ירון מרגולין
- האסכולה הונציאנית (ציור) חלק א' מאת ירון מרגולין
- ולנטין דה בולון צייר גאון – ירון מרגלין בפתח התערוכה בפריז של ולנטין דה בולון
- אן-לואי ז'ירודה דה רוסי-טריוזון
- ברתולומאיוס שפרנגר
- תודעה שלילית – צל על המחשבה
- Negative Consciousness – A Shadow over Thought
- תזונה: צוואת הרפואה הקדומה שהוזנחה
- בדיקות חדשות יכולות להציל חיי חולי כבד שומני 16 שנים טרם זוהתה המחלה
- שיחות ההחלמה ושיחות בין ידידים – על ההבדלים ביניהן
- והדרך לזכייה בבריאות מחדש
- גמישות היא מצב נפשי – אתגר בזרימה ושינוי – אני מבקש להתגמש
- מיכולסטרול גבוהה – מד
- מגילת רות האלמנה שיצאה ממרחב הנוחות
- יש פתרון כולל לבעיות הקשורות לדימוי עצמי, חוסר בטחון וערך עצמי נמוך – דופמין
- חרדה
- מעגל המוזר – התפתחות ה"אני" בשנה התשיעית לחייו
- איך מורידים לחץ וחרדות באופן טבעי? התמודדות עם סטרס ולחץ נפשי
- טיפול נפשי ללא תרופות – חרדות, פחדים וייאוש – תסמינים שיש להם פתרון טבעי, מדויק ומהיר
- למה אני לא מצליח להתמיד – והסוכר
כתיבת תגובה