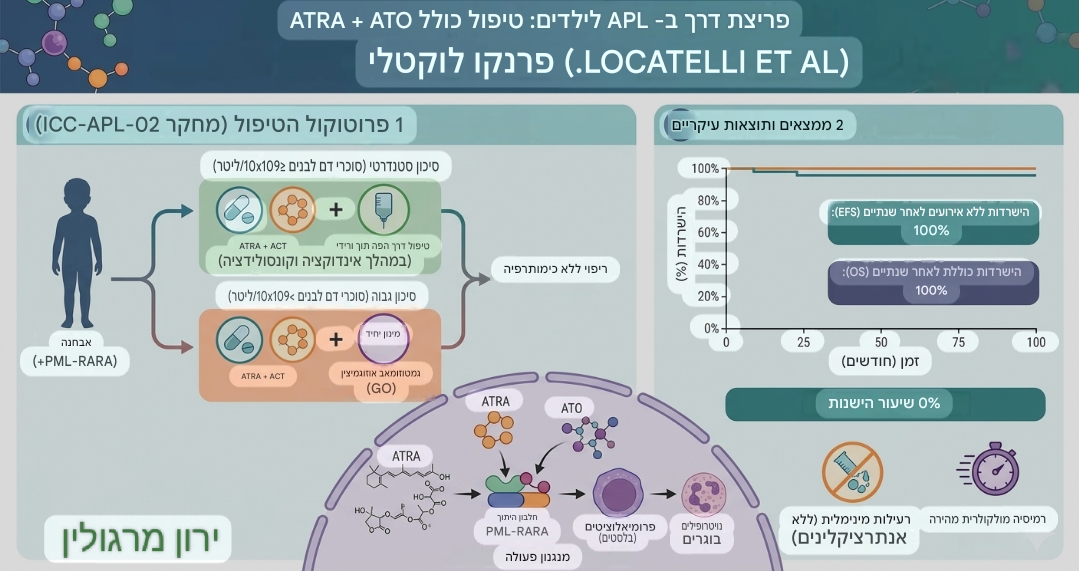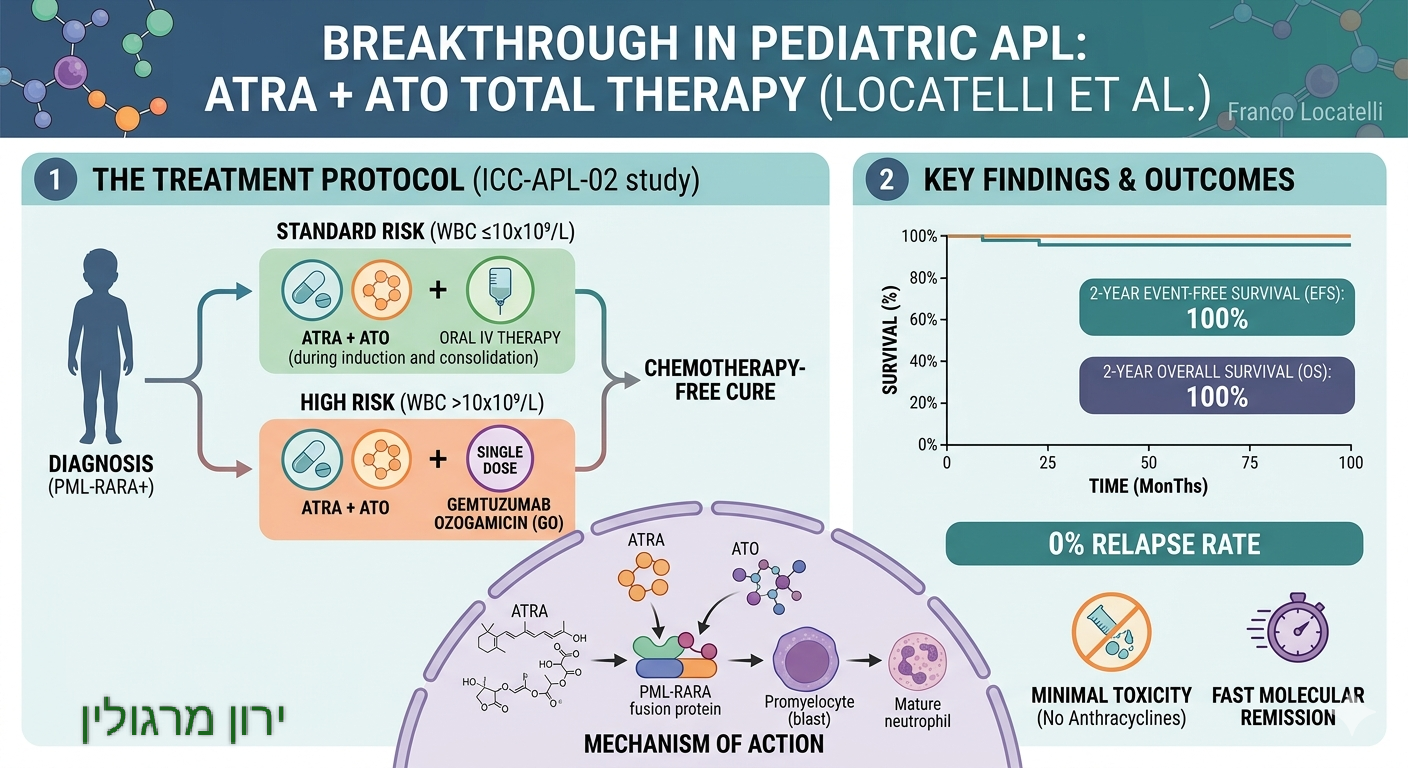
חלק ג' – יציג בהמשך הדברים גורמים טבעיים שנחקרו כתומכים בסביבה תאית שעשויה להפחית את שטף הנזק ולסייע למנגנוני התחזוקה והתיקון.
איור ירון מרגולין.
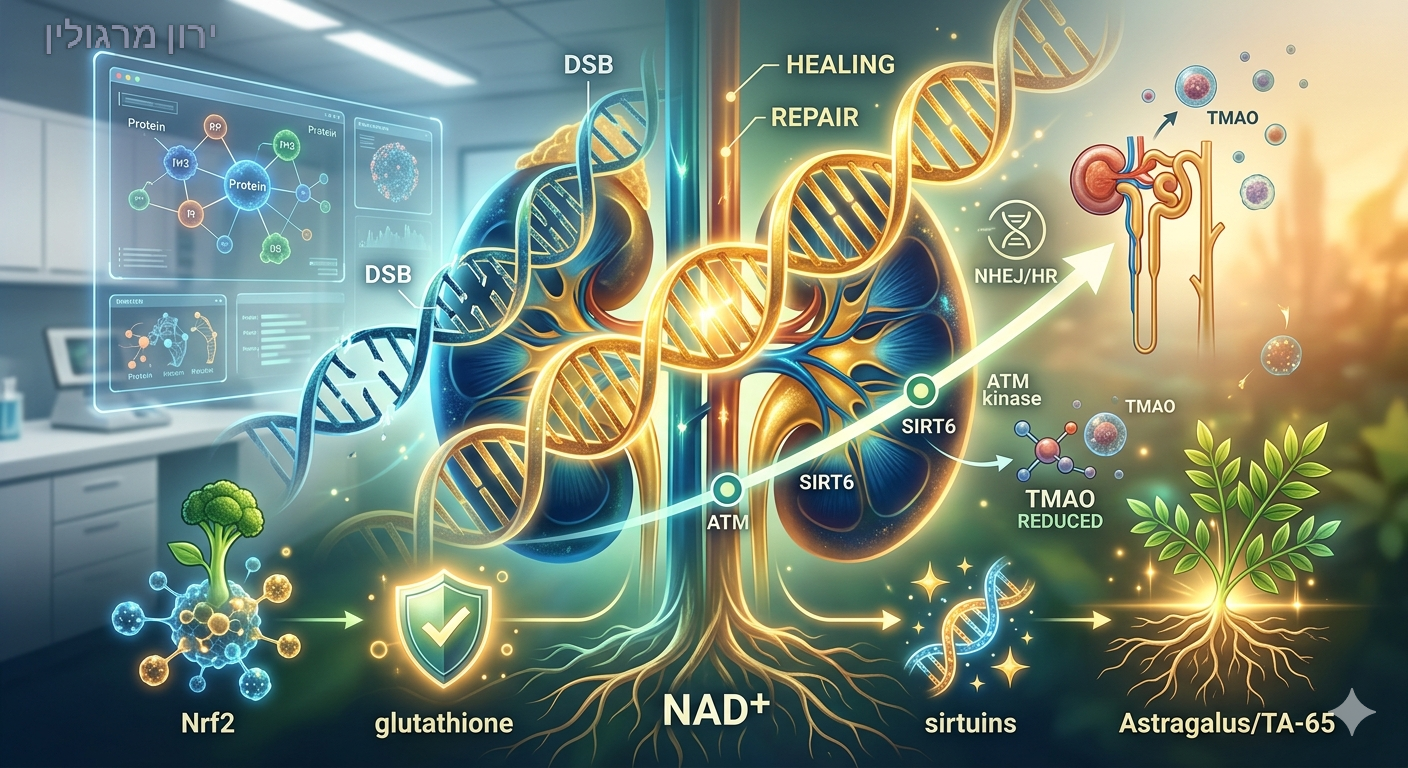
בצד שמאל (הנזק והשבר):
-DNA מוצג בגווני אדום וכתום עם שבר דו–גדילי (DSB) ברור, מוקף בענני עקה חמצונית ורעלנים המייצגים את קריסת המערכת.
בצד ימין (ההחלמה והאפיגנטיקה): ה-DNA משתקם והופך לירוק וכחול זוהר, עטוף ברשתות הגנה מוזהבות של אנזימי התיקון, ה-Sirtuins והאנרגיה המיטוכונדריאלית התקינה
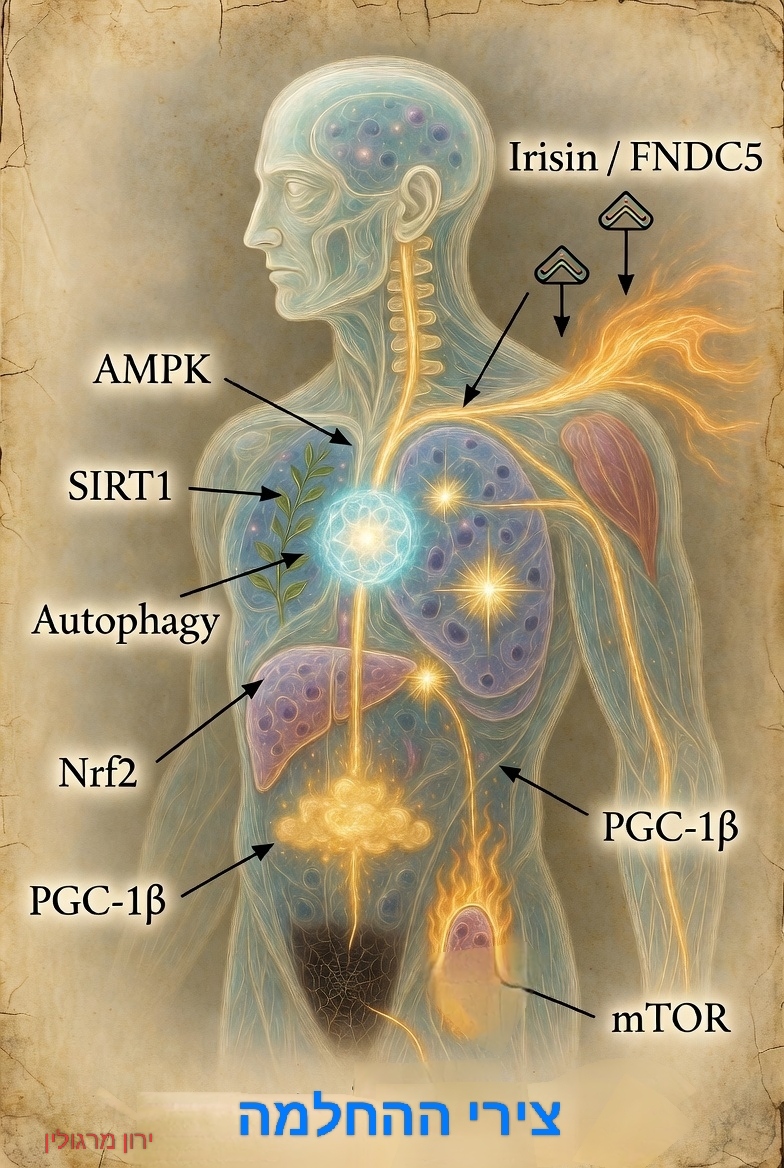
מבוא לחלק ג': מעבר מרפואת מחלות לרפואת חיוניות וצירי החלמה
החלק השלישי של המאמר מציע פתרונות בשני מישורים: הפחתת כוחות ההרס מחוץ לתא (סולפורפאן, Nrf2 חמצון) ותמיכה ישירה במנגנוני הבקרה והאנרגיה בפנים (אסטרגלוס, סירטואינים, NAD⁺).
הגישה הטיפולית המוצעת עוברת מטיפול נקודתי במחלות לשיקום "נתיבי החלמה" פנימיים.
מחלות כליות וסוכרת קשורות למנגנונים משותפים כמו דלקת ועקה חמצונית, המפעילים את ה-DNA ומערכות התיקון (כמוצג במאמרי מפת מסלולי ההחלמה – 3 צירים – מבט כללי). החלק הזה מתמקד בהפעלת צירי הגנה וניקוי (כמו SIRT6 🫐) באמצעות אורח חיים ותזונה🍎, ומדגיש כיצד תאים בוחרים בין תיקון והתחדשות לבין קריסה ופיברוזיס.
רוב הרפואה המודרנית מאורגנת סביב מחלות: מחלת כליות, סוכרת, טרשת עורקים, יתר לחץ דם, כולסטרול גבוה, אלצהיימר, אי־ספיקת לב ומחלות רבות נוספות.
אולם כאשר מתבוננים לעומק במסלולי ההחלמה, הם מנגנונים ביולוגיים הפועלים בגוף, מתגלה תמונה חדשה. מחלות רבות, כך מסתבר, חולקות מסלולים משותפים: דלקת כרונית, עקה חמצונית, פגיעה במיטוכונדריה, נוקשות עורקים, הצטברות שומן ברקמות שאינן מיועדות לכך, פגיעה באיזון ההורמונלי, שבירה דו-גדילית של החומר הגנטי בתא (DSB), וירידה ביכולת התאים להתחדש.
שיקום, ניקוי והפעלת "נתיבי החלמה" משותף לכן, לרוב המחלות, מדובר למעשה בתהליך חדש, גישה אחרת, ממוקד שנועדה לחדש, לטהר ולהחזיר מערכות, מבנים או תהליכים אישיים שנחסמו עם הזמן למצב תקין. בין אם מדובר בשיקום פיזי לאחר נזק, כמו בשבירה דו-גדילית (DSB), טיהור סביבתי או ניקוי מנטלי, השלבים דורשים פעולה שיטתית [מקור1, מקור2, מקור3]. חלק מהדרכים מגיעות מן העבר הרחוק, כשהיום יש לנו הבנה לסיבה שחומרי החלמה פשוטים אלו כל כך הצליחו, כמו קרני שמש, והליכה או מרק עדשים ואפונה …
דוגמאות לצירי החלמה
- הגברת סינתזת גלוטתיון והגנה מפני עקה חמצונית.
- הפעלת Nrf2 והגברת ייצור אנזימי ההגנה של התא.
- שיפור תפקוד המיטוכונדריה וייצור האנרגיה.
- הפעלת AMPK ושיפור גמישות המטבוליזם.
- הפעלת SIRT1 ומנגנוני הסתגלות תאיים.
- הגברת ביטוי Klotho והאטת תהליכי הזדקנות תאית.
- שיקום גמישות כלי הדם.
- הפחתת דלקת כרונית מתמשכת.
עוד על כך במאמרי מפת מסלולי ההחלמה – 3 צירים – מבט כללי

הרפואה הנוכחית להוטה לסייע, היא מוכנה להפוך עולמות כדי להציל חיי אדם, אנשיה נשבעים את שבועת היפוקרטס [מקור]. יש לה רצון טוב, אבל היא בנויה על "זיהוי מחלה" ולא על "מדידת חיוניות". לרופא קל למדוד HbA1c (סוכר) כי יש תרופה שתוריד אותו. מה הוא יעשה עם מדד שמפעיל ציר החלמה?
אין "תרופת מדף" ל-PGC–1α (מלבד אימון גופני, הליכה, שחיה, וקור), אז הציפייה מהרופא שילמד פילוס6, שישנה את צורת מחשבתו, רק כדי שאדם יחלים ממחלה שעלולים למות ממנה לפעמים נשמעת כמו זעקה בימי הביניים.
מה גורם לתא להישאר תא בר־תיקון, ומה דוחף אותו לעבור מן השבר אל תהליך "בלאי" ביולוגי: סנסנס, פיברוזיס וקריסה?

חלק ג' יתמקד תחילה בשאלה:
כיצד גבול ההגנה של הכרומוזום קובע אם התא יישאר במסלול של תיקון והתחדשות — או ייסחף לתהליך "בלאי" ביולוגי: סנסנס, פיברוזיס ואובדן תפקוד. וכן
מה ניתן לעשות כדי להפעיל את צירי ההגנה🛡️, ההחלמה וניקוי 🧹 במקרה זה של שבר דו־גדילי, אציין גם מה ניתן לעשות נוכח ה DSB ❓
כאשר שבר דו־גדילי (DSB) נובע משחיקה טלומרית, התא עובר מתהליכי תיקון והתחדשות למסלול של "בלאי" ביולוגי: סנסנס, פיברוזיס ואובדן תפקוד. הטלומרים, רצפים חוזרניים בקצות הכרומוזומים (כמו TTAGGG), משמשים כ"פקק" הגנה חיוני. הם מונעים מקצות הכרומוזומים להיתפס כשברים גנומיים המצריכים תיקון, מה שעלול להוביל לאיחוי שגוי ומסוכן של כרומוזומים.
קומפלקס חלבוני ה-Shelterin (כולל TRF1, TRF2, POT1 ועוד) מגן על הטלומרים ומארגן אותם במבנה מוגן (T-loop). כאשר הגנה זו נשחקת (בשל עקה חמצונית, דלקת, או חלוקה מרובה), קצה הכרומוזום נחשף ומזוהה בטעות כשבר, מה שגורם לתגובת נזק כרונית ל-DNA (DDR).
ההגנה נשחקת, ונוצרת אי-יציבות כרומוזומלית דרך מנגנון NHEJ, המקדם סנסנס תאי (הזדקנות) והפרשת גורמים דלקתיים (SASP). ברקמות, ובמיוחד בכליות, שחיקה טלומרית זו היא מנוע דלקתי-פיברוטי, המוביל להצטלקות, לאובדן התחדשות ולירידה בתפקוד הכלייתי.
כאשר הפגיעה הגנומית DSB נובעת משחיקה טלומרית
הטלומרים ממלאים תפקיד שנראה במבט ראשון פשוט — הגנה על קצה הכרומוזום מפני שחיקה ואובדן מידע גנטי בעת חלוקת התא. אולם מבחינה ביולוגית, תפקידם עמוק בהרבה. קצה כרומוזום חשוף דומה במידה מטרידה לשבר דו־גדילי (DSB) ב־DNA: בשני המצבים קיימת בקצה DNA חופשיות, שעלולה להיקלט על־ידי מערכות ניטור הנזק של התא כאות מצוקה המחייב גיוס של מסלולי תיקון🛠️. מכאן נובעת אחת הבעיות העקרוניות ביותר בתחזוקת הגנום:
כיצד התא מבחין בין שבר אמיתי שיש לתקנו במהירות, לבין קצה כרומוזום תקין שאותו דווקא אסור "לתקן".
לשם כך פיתח התא מערכת הגנה ייעודית לטלומרים. רצפי הטלומר החוזרניים, יחד עם קומפלקס חלבונים ייחודי, מעצבים את קצה הכרומוזום כך שלא יזוהה בטעות כ־DSB.
במצב תקין, קצה הכרומוזום "מוסתר" ממערכות התיקון; הוא מוגן מפני הפעלת מסלולי חירום של DNA damage response, ומפני איחוי לא־מבוקר עם קצה כרומוזומלי אחר. כך, הטלומר אינו רק "פקק" מכני בקצה הכרומוזום, אלא גבול ביולוגי פעיל המונע מהתא להפנות את מנגנוני התיקון אל מקום שבו תיקון יהיה למעשה שגיאה.
בנקודה זו מתברר עד כמה טלומרים ושברי DNA קשורים זה בזה. מסלולי תיקון כמו NHEJ נועדו להציל את הגנום כאשר שבר דו–גדילי (DSB) מתרחש, אך אם אותם מסלולים יופעלו על קצות כרומוזומים לא־מוגנים, התוצאה עלולה להיות הרסנית: איחוי קצה־לקצה בין כרומוזומים, יצירת כרומוזומים דיצנטריים, קריעה מחודשת בזמן חלוקה, ומעגל של אי־יציבות כרומוזומלית ההולך ומעמיק. במילים אחרות, אחת ממשימותיו החשובות ביותר של התא איננה רק לתקן DSB, אלא גם למנוע ממנגנוני התיקון לגעת בטלומר כשהוא תקין. מכאן שטלומרים אינם נושא צדדי בדיון על נזקי DNA; הם חלק מן ההבחנה הבסיסית בין תיקון נכון לבין תיקון שגוי.
הטלומר כמבנה מוגן: מדוע קצה כרומוזום אינו אמור להיתפס כשבר
הטלומרים בנויים ברוב התאים האאוקריוטיים מרצפים חוזרניים עשירים בגואנין, אשר אצל האדם מורכבים מחזרות של הרצף TTAGGG. אך רצף זה לבדו אינו מספיק. ההגנה הטלומרית נשענת גם על קומפלקס חלבונים ייחודי, הידוע בשם shelterin, הכולל בין השאר את TRF1, TRF2, POT1, TIN2, TPP1 ו־RAP1. קומפלקס זה מארגן את קצה הכרומוזום במבנה תלת־ממדי מוגן, ולעיתים גם בלולאה טלומרית (T-loop), המסייעת "להחביא" את קצה ה־DNA החד־גדילי והדו־גדילי מפני מערכות החישה של נזקי DNA.
זהו מנגנון חיוני במיוחד משום שמערכות החישה הגנומיות פועלות מתוך היגיון הישרדותי אגרסיבי: קצה DNA חופשי נתפס כאיום. חיישנים מולקולריים כמו ATM, ATR, קומפלקס MRN וחלבוני בקרה נוספים נועדו לזהות במהירות מבנים החשודים כשבר, לגייס פוספורילציה של H2AX, להפעיל מחסומי מחזור תא ולקרוא לחלבוני תיקון.
כל עוד shelterin מתפקד היטב, הטלומר מוגן מפני הפעלה שגויה של מערכות אלה.
אך כאשר ההגנה הטלומרית נחלשת — בשל קיצור טלומרים, עקה חמצונית, שינויים אפיגנטיים, דלקת כרונית או ירידה בתפקוד החלבונים המגנים — קצה הכרומוזום עלול להפוך בעיני התא ל"שבר". ברגע זה, הגבול בין תחזוקת כרומוזום תקינה לבין תגובת חירום גנומית מתחיל להיטשטש.
TRF2, למשל, נחשב לאחד השומרים המרכזיים מפני הפעלת תגובת ATM ואיחוי טלומרים דרך NHEJ, בעוד POT1 מסייע להסתיר את ה־single-stranded overhang ולמנוע הפעלה של מסלולים התלויים ב־ATR. במילים אחרות, ההגנה הטלומרית איננה פסיבית: היא מנטרלת באופן אקטיבי את מנגנוני זיהוי הנזק במקום שבו עצם ההפעלה שלהם תהיה מזיקה. כאן טמון פרדוקס עמוק של הביולוגיה התאית — אותם מסלולי תיקון המצילים את התא משבר דו–גדילי (DSB) אמיתי, עלולים להפוך לגורם הרס אם יופעלו על קצה כרומוזום שאיבד את מעטפת ההגנה שלו.
הערה: TRF2 הוא חלבון חיוני בגוף האדם המגן על קצות הכרומוזומים (הטלומרים) מפני נזק. TRF2 הוא מרכיב מרכזי בקומפלקס חלבונים בשם Shelterin. תפקידו העיקרי הוא להגן על קצות הכרומוזומים כדי שלא יזוהו בטעות על ידי מערכות התיקון של התא כנזק לחומר הגנטי ל-ל-DNA [מקור].
איור ירון מרגולין
כאשר ההגנה נשחקת: מן הטלומר המקוצר אל תגובת נזק כרונית
הטלומרים מתקצרים בהדרגה עם חלוקות תאיות חוזרות, בין היתר בשל "בעיית סוף השכפול" (end-replication problem), שבגללה מנגנון השכפול מתקשה להשלים במלואו את קצה הגדיל הליניארי. אך
השחיקה הטלומרית איננה תוצאה של חלוקה בלבד.
עקה חמצונית, דלקת כרונית, רעלנים מטבוליים, פגיעה מיטוכונדריאלית והצטברות ROS על כולם אורח חיינו משפיע וכשבעיקר התזונה שלנו מפעילה מנגנונן הרס אלו, הם מאיצים את הנזק הטלומרי, לעיתים גם ללא חלוקה רבה.
טלומרים רגישים במיוחד לנזק חמצוני, בין השאר בשל עושרם בגואנין, בסיס הנוטה לחמצון, ובשל הקושי היחסי לתקן נזקי DNA באזורי קצה כרומוזומליים דחוסים.
כאשר הטלומר מתקצר או מאבד את ארגון ה־shelterin שלו, התא עלול להפעיל סביבו תגובת נזק ל־DNA.
מופיעים telomere dysfunction-induced foci, חלבוני DDR מגויסים לאזור, ונתיבי p53/p21 או p16^INK4a/Rb עשויים להוביל לעצירת מחזור ממושכת.
במצבים מסוימים זו עשויה להיות תגובת הגנה חיונית: מנגנון שנועד למנוע חלוקה של תא בעל גנום בלתי יציב. אך ברקמות שנפגעו כרוניות, ובמיוחד ברקמות שבהן פגיעה חוזרת, מצטברת לאורך שנים, כמו באובדן גמישות העורקים, יתר לחץ דם, אי ספיקת כליות, סוכרת, מחלות לבביות, אלצהיימר תגובה זו עלולה להפוך למצב מתמשך של סנסנס תאי.
המשמעות של סנסנס איננה רק "עצירת חלוקה". תא סנסנטי נשאר חי, אך משנה את זהותו התפקודית. תא סנסנטי (מזדקן) נעשה למפעל של הפקת רעלים: הוא מתחיל להפריש ציטוקינים, כימוקינים, פרוטאזות וגורמי גדילה במכלול המוכר כ־SASP. הפרשה זו עשויה להחריף דלקת, לפגוע בתאים שכנים, לעודד הפעלה של פיברובלסטים ולהזין מעגל של שיפוץ פתולוגי ברקמה. לכן, שחיקת טלומרים אינה רק בעיה של "שעון ביולוגי" או של ירידה בכושר החלוקה; היא עלולה להפוך מנזק גנומי נקודתי למנוע דלקתי־פיברוטי מתמשך.
טלומרים, NHEJ ואי־יציבות כרומוזומלית: כשהתיקון עצמו הופך לאיום
אחד ההיבטים החשובים ביותר של טלומרים בהקשר של DSB הוא הסכנה שבתיקון שגוי. כאשר קצה כרומוזום מאבד את ההגנה שלו, התא עלול לזהות אותו כשבר דו–גדילי (DSB) ולגייס מסלולי חיבור קצוות — בראש ובראשונה NHEJ. אלא שבמקרה זה אין מדובר בשבר "אמיתי" שיש לאחותו עם בן־זוגו המקורי, אלא בקצה כרומוזומלי שצריך היה להישאר מבודד. אם הוא מתאחה עם כרומוזום אחר, נוצר מבנה כרומוזומלי בלתי תקין, לעיתים כרומוזום דיצנטרי, שמתקשה להיפרד כראוי בזמן מיטוזה. התוצאה עלולה להיות מעגל breakage-fusion-bridge: הכרומוזומים נקרעים מחדש, מתאחים שוב, ויוצרים אי־יציבות גנומית מתקדמת.
מנקודת מבט אונקולוגית, חקר מחלת הסרטן, תהליך כזה עשוי להוות קרקע להצטברות שינויים כרומוזומליים, אמפליפיקציות, מחיקות ומוטציות המקדמות טרנספורמציה ממארת. אולם בהקשר של מחלת כליות כרונית, החשיבות רחבה יותר. גם אם תא פגוע לא יתקדם למחלת הסרטן, עצם ההפעלה המתמשכת של תגובת נזק, עצירת המחזור, הסנסנס והפרשת ה־SASP עשויות להספיק כדי לשנות את גורל הרקמה. בכליה, שבה תאי אפיתל טובולריים, תאי אנדותל ותאי אינטרסטיציום חשופים לעקה חוזרת, אובדן ההגנה הטלומרית עלול לתרום לא רק לאי־יציבות גנומית, אלא גם להצטלקות, לאובדן כושר התחדשות ולירידה מתמשכת בתפקוד הכליה ולמנוע טיהור דם מרעלים שהולכים ומצטברים בגוף.
כאן מתחדד שוב המסר המרכזי של המאמר: לא כל כשל בתיקון DNA מוביל לסרטן, אך
כמעט כל כשל מתמשך בתחזוקת הגנום עלול לדחוף את התא למסלול פתולוגי.
לעיתים המסלול הזה הוא ממאיר; לעיתים הוא פיברוטי; ולעיתים הוא פשוט מסלול של שחיקה איטית — תא שכבר אינו מתחדש, אך ממשיך להפריש אותות נזק ולפגוע בסביבתו מקדם תחלואה.
בהקשר הפיזיולוגי, נפרון מייצג יחידה תפקודית המסננת דם, סופגת מחדש את האלקטרוליטים המסוננים, המומסים והנוזלים, ומפרישה פסולת ואלקטרוליטים מיותרים ומים [מקור1, מקור2, מקור3, מקור4, מקור5].
הגלומרולוס (פקעית) מבשרת את תחילת הנפרון, אבל היא עצמה חלק ממערכת הדם. היא נמצא בתוך מבנה סגור דמוי קערה או משפך סגור שנמצא בתחילתו של כל נפרון, מבנה זה מכונה: כמוסה גלומרולרית (a glomerular capsule) אשר ידוע כקופסית באומן, או הקופסית על שם באומן שבאיור למעלה (האיור לקוח מויקיפדיה).
לנימים שמרכיבים את הפקעית נקבוביות קטנות בדפנות, מכאן אנשים רבים מכנים אותם המסננת. רוב הנימים האלו דחוסות בין העורקים והוורידים (הכלים הקטנים המעבירים דם אל הנימים ואוספים בהמשך גם דם מהנימים, לאחר שנוקה אל זרם הדם), לעורקים שמובילים את הדם אל הנימים, שבתוך קופסת באומן קוראים העורקים האפרנטיים [(AA) האפרנטיים (the efferent arterioles] [מקור]. האיור למטה מראה את העורקים האפרנטיים –
הנימים הגלומרולריים, שמרכיבים את הפקעית, שמקורם כאמור לעיל מהעורקים האפרנטיים, מיוחדים בכך שהם מסוגלים לעמוד בלחצים הידרוסטטיים גבוהים, שמהווים כוח מניע עיקרי לסינון שמתרחש בפקעית שבקופסית באומן [מקור]. תאים אלו עומדים בלחצים הידרוסטטיים גבוהים, שמהווים גורם סיכון מכני קבוע.
השילוב הייחודי הזה בין הלחץ ההידרוסטטי הגבוה בפקעית הכליה (הגלומרולוס) לבין ריכוז הרעלנים הגבוה בדם, הוא שיוצר פצצה מולקולרית מתקתקת בתאי האנדותל. הלחץ המכני הכרוני מותח את דפנות התאים ומפעיל אנזימים (כמו NADPH oxidase) שמייצרים שטף אדיר של רדיקלים חופשיים (ROS) מקומיים. במקביל, רעלנים המצטברים בדם פוגעים פיזית בדופן התא ומנטרלים את נוגדי החמצון הטבעיים שלו.
שטף ה-ROS המקומי הזה נודד ישירות לגרעין ותוקף את הטלומרים. הסיבה לכך היא שהטלומרים עשירים ברצפים חוזרים של הבסיס חנקני גואנין (Guanine), שהוא הרכיב הרגיש והפגיע ביותר בגנום לחמצון. רדיקלים חופשיים מחמצנים את הגואנין והופכים אותו למולקולה פגומה בשם 8-oxoG.
ברגע שנוצר ה-8-oxoG בטלומר, קומפלקס ה-Shelterin (המגן הטבעי שעליו דיברנו) אינו מצליח עוד להיקשר ל-DNA והוא קורס. התא מפרש את קריסת ה-Shelterin ואת קצה הכרומוזום החשוף והמחומצן כאילו מדובר בשבר דו–גדילי (DSB) קטסטרופלי. האזעקה המולקולרית מופעלת, התא עוצר את חלוקתו ומדרדר במהירות למצב של סנסנציה (הזדקנות תאית) והפרשת רעלים (SASP) המזינים את תהליך הפיברוזיס בכליה.
טלומרים בכליה: מן ההזדקנות התאית אל אובדן נפרונים ופיברוזיס
הכליה היא אחת הרקמות שבהן המתח בין הישרדות, תיקון והתחדשות בולט במיוחד. תאי האבובית הפרוקסימלית, מכונה גם מקורבת למשל, פועלים בסביבה עתירת חמצון ומיטוכונדריות, צורכים כמויות גדולות של ATP, וחשופים לרעלנים, להיפוקסיה, לעומס מטבולי ולשינויים המודינמיים. בתנאים כאלה, גם פגיעה טלומרית מתונה עלולה לקבל משמעות מצטברת. כאשר תא טובולרי נכנס לתהליך "בלאי" ביולוגי-סנסנס, הוא אינו רק "יוצא מן המשחק"; הוא עלול להפוך למוקד הפרשה של גורמים פרו־דלקתיים ופרו־פיברוטיים, לשבש את המיקרו־סביבה המקומית ולעכב תיקון תקין של האפיתל לאחר פגיעה.
מחקרים בעשור האחרון קשרו בין קיצור טלומרים, ביטוי מוגבר של סמני סנסנס ופעילות DDR (לבין ירידה בתפקוד (הזדקנות כלייתית, התקדמות CKD ופגיעה בהתאוששות לאחר AKI. אמנם הקשרים אינם אחידים בכל המצבים, והטלומר אינו ההסבר היחיד או הבלעדי לירידה תפקודית (הזדקנות) הכליה, אך התמונה המצטברת עקבית למדי: ככל שההגנה הטלומרית נשחקת, כך גדל הסיכון לכך שתאים כלייתיים יעברו ממצב של תיקון אדפטיבי למצב של עצירת מחזור כרונית, ירידה בכושר ההתחדשות והזנת תהליכי פיברוזיס. במילים אחרות, הטלומר איננו רק מדד לזמן שעבר על התא; הוא עשוי להיות אחד הגורמים הקובעים אם פגיעה כלייתית תסתיים בהחלמה ואפילו בהחלמה חלקית — או בצלקת קבועה, דיאליזה והשלכותיה.
יש לכך משמעות עמוקה במיוחד בהקשר של CKD. במחלה כרונית, התא הכלייתי אינו ניצב מול אירוע חד־פעמי אחד, אלא מול רצף של פגיעות חוזרות: ROS, דלקת, רעלנים אורמיים, הפרעות מטבוליות, הפרעות בכלי הדם והפרעה באיכות המיטוכונדריות. כל אחד מן הגורמים הללו עלול לפגוע בטלומרים ישירות או בעקיפין, וכל פגיעה כזו מקטינה מעט את מרחב הפעולה של התא. במוקדם או במאוחר, חלק מן התאים חוצים סף שממנו קשה פזהותם, הם עוברים נזק של שינוי זהות: תא שהיה אמור להשתקם הופך לתא חלש, נעדר יכולות תפקודיות (מזדקן), ותא מוחלש הופך לגורם פעיל בהתקדמות המחלה.
החשיבות של הטלומרים בדיון על DSB נובעת אפוא משני כיוונים משלימים. מצד אחד, הם מגינים על קצה הכרומוזום מפני זיהוי שגוי כשבר דו־גדילי (DSB), ובכך מונעים הפעלה מזיקה של מסלולי תיקון. מצד אחר, כאשר הגנה זו נשחקת, הטלומר עצמו הופך למוקד של תגובת נזק כרונית, סנסנס, ולעיתים אי־יציבות כרומוזומלית. בכך הוא מחבר בין תחזוקת הגנום לבין השאלה הרחבה יותר של גורל התא: האם התא יוכל להמשיך לתקן, להתחלק ולהתחדש, או שיעבור למצב של עצירה, הפרשת אותות נזק ותרומה פעילה לפיברוזיס.

טלומרים כציר מעבר בין נזק גנומי לבין גורל התא
אם התא הכלייתי נמצא כבר תחת שטף נזק מתמשך, השאלה המעשית בשלב זה היא אחת: כיצד להפחית את עומס הפגיעה, לשמר אנרגיה, להגן על הטלומרים, לתמוך בגלוטתיון וב־NRF2, ולחזק את הסביבה שבה מנגנוני התיקון פועלים.
כיצד להפחית את שטף הנזק ולתמוך בתא בר־תיקון כאשר הפגיעה הגנומית DSB נובעת משחיקה טלומרית
מכאן מתחדדת גם השאלה המעשית. אם פגיעה טלומרית איננה רק סמן של זמן ביולוגי, אלא מנגנון המסוגל להפעיל תגובת נזק כרונית, לדחוף את התא לתהליך "בלאי" ביולוגי – סנסנס ולתרום לפיברוזיס כלייתי, אין די בתיאור המבנה הטלומרי ובמסלולי הנזק הכרוכים בשחיקתו. יש לשאול גם
האם ניתן להפחית את הכוחות המאיצים את הפגיעה בקצות הכרומוזומים, ובכך לשמר תא שעדיין מסוגל לבחור בתיקון ובהישרדות במקום בעצירת מחזור ובהפרשת אותות נזק.
שאלה זו מחייבת זהירות. אין כיום דרך פשוטה, מבוססת ובטוחה "לתקן טלומרים" במובן הישיר, ובוודאי לא להפוך בקלות טלומר שנשחק לטלומר צעיר ומתפקד. אך דווקא משום כך, הדיון המעשי חייב להתמקד לא בהבטחות של הארכה מלאכותית, אלא בצעד הצנוע והחשוב יותר: 1. התבוננות במתרחש מחוץ לקליניקה הממסדית – נושא מודר מהספרות המחקרית, עליו כתבתי כאן. 2. הפחתת שטף הנזק המופנה אל הטלומרים, צמצום הסביבה הדלקתית והחמצונית השוחקת אותם, ותמיכה במערכות התחזוקה התאיות שעשויות להרחיק את התא מן הנקודה שבה קצה כרומוזום פגוע מתחיל להיתפס כשבר דו־גדילי (DSB).
מכאן ואילך, אפוא, המוקד איננו "מה מאריך טלומרים" במובן הפשטני, אלא מה עשוי להפחית את הסיכוי שפגיעה טלומרית תהפוך לאות נזק כרוני, לתהליך "בלאי" ביולוגי: סנסנס ולפיברוזיס (צלקת). זהו מעבר מן השאלה כיצד טלומרים נפגעים — אל השאלה כיצד ניתן, בזהירות ותוך הקשבה עמוקה לתוצאות המחקר המדעי והימנעות מהבטחות יתר, או סיכון, או הפחתה בשימוש תרופתי, שהושקע בו הרבה כסף. כלומר מבלי לגרום נזק לתעשייה. אבל, לתמוך בתא בר־תיקון ככל האפשר, בעזרת הפעלתם של צירי תיקון כאשר מקור האיום הגנומי מצוי בקצות הכרומוזומים עצמם.
מה ניתן לעשות בפועל כאשר הנזק לחומר הגנטי קשור לטלומרים
כאשר מקור חלק מן הנזק הגנומי הוא שחיקה טלומרית, המטרה אינה "להאריך טלומרים" באופן מלאכותי, אלא להפחית את הכוחות השוחקים אותם ולשמר תא שעדיין מסוגל לבחור בתיקון ולא בסנסנס.
1) להפחית עקה חמצונית כרונית
טלומרים רגישים במיוחד ל־ROS, ולכן זהו הצעד הראשון.
בפועל זה אומר:
- סולפורפאן ממקור של נבטי ברוקולי / ברוקולי צעיר / ירקות מצליבים
- תזונה שמפחיתה דלקת ועומס חמצוני
- הימנעות מחשיפה מיותרת לרעלנים, עישון, מזון אולטרה־מעובד ועומס גליקמי חריף
2) לשפר את סביבת החיים של התא, לא רק את ה־DNA
פגיעה טלומרית מואצת כאשר התא חי בסביבה של דלקת, שינה גרועה, עקה מטבולית ופגיעה מיטוכונדריאלית.
לכן יש היגיון ב:
- חשיפה סדירה לאור יום / שמש בוקר
- שינה מספקת וסדירה
- צמצום סטרס ביולוגי כרוני ככל האפשר
3) לתמוך בצירי עמידות תאית ולא רק "לכבות נזק"
כאן נכנסים תוספי תזונה שמפעילים צירי החלמה:
- רסברטרול — לא כ"מתקן טלומרים", אלא כחומר שנחקר בהקשר של SIRT1, עקה חמצונית, דלקת והזדקנות תאית
- בהמשך המאמר, תחת SIRT1/SIRT6, נכון נחזור אל קמח זרעי הענבים ונמקם אותו בצורה מדויקת יותר
4) לשמור על סביבה מטבולית שאינה שוחקת טלומרים
כלומר:
- להפחית היפרגליקמיה, גליקציה, ליפוטוקסיות ודלקת כרונית
- להעדיף תזונה פשוטה, נקייה, עשירה בירקות, עם פחות עומס סוכר ומזון מעובד
- לדאוג שלא יהיו חסרים תזונתיים משמעותיים הפוגעים בגלוטתיון, במתילציה ובמיטוכונדריה
5) להבין את גבול הידע
המסר המדעי החשוב הוא זה:
כאשר הנזק לחומר הגנטי נובע מטלומרים, יש מקום לשתי אסטרטגיות משלימות — האחת להפחית את הכוחות השוחקים את הטלומר, עליה דנתי למעלה, והרמה השנייה שמעניינת לדעתי יותר, ותובא להלן, היא לבחון בזהירות התערבויות שנחקרו כמשפיעות על תחזוקת טלומרים או על טלומראז, ובראשן אסטרגלוס (קדד קרומי) ונגזרותיו (נושא שרק בשנים האחרונות שב והוכח מחקרית [מקור]) [מקור]. עם זאת, כל ניסיון "להפעיל טלומראז" חייב להיבחן גם לאור הסיכון התאורטי של תמיכה בתאים טרום־ממאירים או ממאירים, שכן טלומראז הוא גם אחד המנגנונים המרכזיים המאפשרים לתאי סרטן לחמוק מהזדקנות וממוות. וכאמור ללכת על קצה האצבעות כדי לא לפגוע בבינה המלאכותית, ובתעשיית התרופות וגרוע מכך במיליוני מהדהדי מסריה.


האם ניתן לתמוך בטלומרים גם באמצעות תוספים — ומה המשמעות של זה בכליה פגועה?
לצד העקה החמצונית, הדלקת הכרונית, הפגיעה המיטוכונדריאלית והעומס המטבולי המאיצים שחיקה טלומרית, נבדקו גם התערבויות שמטרתן לתמוך ישירות בתחזוקת הטלומרים עצמם. אחת הידועות שבהן היא TA-65, נגזרת של Astragalus membranaceus (אסטרגלוס), הנחשבת למפעילת טלומראז. בעבר היה מקום לספק אם קיימת לגביה ראיה אנושית מספקת, אך כיום התמונה מורכבת יותר: במספר מחקרים קליניים, ובכללם מחקר אקראי מבוקר, וכן במטא־אנליזה עדכנית, נמצאה השפעה אפשרית של TA-65 על אורך הטלומרים בלויקוציטים, ובחלק מן העבודות אף הודגמה האטה בקיצורם או הארכה מתונה. גם ויטמין D₃ התגלה בשנים האחרונות כמועמד מעניין יותר מכפי שנהוג היה לחשוב: ממצאי החוקרים מלמדים כי ייתכן שניתן להשפיע, במידה מסוימת, על אורך הטלומרים או על קצב שחיקתם בעזרת קרני 🌞 שמש [מקור]. אולם השאלה החשובה בכליה איננה אם טלומרים בדם התארכו בכמה עשרות זוגות בסיס, אלא באילו תנאים חי התא הכלייתי עצמו.
בכליה, ובייחוד בפקעית, הטלומרים אינם נשחקים בחלל ריק. הם מצויים בתוך רקמה הנתונה לעיתים ללחץ תוך־גלומרולרי גבוה, להיפרפילטרציה, לעודף חלבון בלתי מותאם, לרעלנים אורמיים, לחומצת שתן, לתוצרי גליקציה, ל־TMAO, לפגיעה מיטוכונדריאלית ולשטף ROS. לכן גם אם חומר כלשהו מסוגל לעורר טלומראז או להאט קיצור טלומרים בלויקוציטים, אין בכך עדיין הוכחה שהוא מגן על הפקעית או משקם תא אנדותל, פודוציט או תא אבוביתי החשופים מדי יום לעומס מכני, מטבולי וחמצוני.
במילים אחרות, הארכת טלומרים בדם איננה שקולה לשיקום הטלומרים בכליה, ובוודאי לא להחלמה של רקמה שכבר מצויה במסלול של תהליך "בלאי" ביולוגי – סנסנס, דלקת ופיברוזיס.
משום כך, אם מבקשים לדבר ביושר על "תמיכה טלומרית" במחלת כליות, יש להבחין בין שתי שאלות שונות.
השאלה הראשונה היא האם ישיבה נוכח קרני השמש ויצירת ויטמין D — מסוגל להשפיע על מדדי טלומרים במחקרים אנושיים. על כך אפשר כיום להשיב בזהירות: ייתכן שכן, לפחות במידה מסוימת.
השאלה השנייה, והחשובה הרבה יותר, היא האם הוא משנה את תנאי השחיקה עצמם בתוך הכליה: האם הוא מפחית ROS, משקם גלוטתיון, מחזק את Nrf2 🥦, מקל על העומס המיטוכונדריאלי, מפחית לחץ תוך־גלומרולרי, מצמצם היפרפילטרציה, מוריד חומצת שתן ומקטין את העומס הרעלי־דלקתי המזין את הפגיעה הטלומרית.
כל עוד התשובה לשאלה השנייה אינה ברורה, אין לראות בתוספים אלה "פתרון לטלומרים", אלא לכל היותר תוספת אפשרית בתוך אסטרטגיה רחבה הרבה יותר. כמו כן יש לקחת בחשבון שהפעלת מסלולי החלמה בעזרת קליפות אננס או קרני שמש מאוד זולים, ומי שמממן את המחקרים עלול להקשות בנושא זה לכאורה.
מכל מקום בהקשר הכלייתי, כדאי להתבונן בתהליכי החלמה שצלחו [מקור1, מקור2, מקור3, מקור4, מקור5, מקור6] המבט על טלומרים חייב להישאר מפוכח. לא די לשאול כיצד לעורר טלומראז או כיצד להאריך טלומרים בדם; צריך לשאול כיצד להוציא את הפקעית עצמה, מסביבת השחיקה שבה היא לכודה.
כיצד להפחית את הלחץ התוך־כלייתי, כיצד לצמצם את ההיפרפילטרציה, כיצד להפחית עומס חלבוני שאינו מותאם, כיצד להוריד TMAO וחומצת שתן, כיצד להקטין את זרם הרעלנים האורמיים, וכיצד לשקם את ההגנה החמצונית והמיטוכונדריאלית של התא.
רק בתוך מסגרת כזו ניתן לדון גם בתוספים בעלי פוטנציאל טלומרי — לא כהבטחת־יתר, אלא כחלק משאלה עמוקה יותר: כיצד משמרים בתא הכלייתי תנאים שבהם הטלומרים אינם נאלצים להישחק מהר מן היכולת של התא לתקן, להתחדש ולהימנע מצלקת.
2) הרמה השנייה — התערבויות שנחקרו בהקשר של תחזוקת טלומרים או הפעלת טלומראז:
- אסטרגלוס / cycloastragenol / TA-65 ודומיו
- אולי בהמשך גם רסברטרול, פעילות גופנית, דפוסי תזונה מסוימים — אבל אסטרגלוס הולך ומוכח על ידי החוקרים כחלק מהציר הישיר יותר כשמדברים על טלומרים עצמם.
יש לקחת בחשבון ככל שהחומר שמקדם החלמה זול יותר, נגיש לכולם, כמו קליפות בצל, זרעי ענבים, נבטי ברוקולי ואפילו הקליפות הקשות של בוטנים וקליפת אננס כך קשה יותר לקדם מחקר יעיל על תרומתו לבריאות ומימון. יחד עם זאת בלעדי מחקר רציני לא ניתקדם אל רפואת העתיד. ובכל זאת מדי פעם, בפריפריה הרחוקה מצליחים חוקרים לפעול ולקדם את האנושות.
מה ניתן לעשות בפועל כאשר הנזק הגנומי נובע משחיקה טלומרית
כאשר הפגיעה בחומר הגנטי נובעת משחיקה טלומרית או מאובדן ההגנה על קצות הכרומוזומים, אפשר לחשוב על שתי שכבות של התערבות. השכבה הראשונה איננה מכוונת לטלומר עצמו, אלא לכוחות המאיצים את שחיקתו: עקה חמצונית, דלקת כרונית, עומס מטבולי ופגיעה מיטוכונדריאלית. במישור זה, יש היגיון בהפחתת ROS, בתמיכה במנוע התיקון המרכזי של הגוף – הגלוטתיון וב־Nrf2 🥦, בשיפור איכות השינה🛌 והחשיפה לאור יום (קרני 🌞), ובהעדפת תזונה המקטינה דלקת ועומס גליקמי [מקור].
לצד גישה זהירה זו, קיימת גם שכבה שנייה, מעניינת יותר: ניסיון להשפיע ישירות על תחזוקת הטלומרים. כאן נכנסים חומרים שנחקרו כמעודדי טלומראז או כתומכים באורך טלומרים, ובראשם נגזרות של Astragalus membranaceus כגון cycloastragenol ו־TA-65. בשנים האחרונות פורסמו מחקרים וניסויים קטנים המצביעים על אפשרות להאריך טלומרים בלויקוציטים או לשפר מדדי טלומרים באמצעות תוספים המבוססים על אסטרגלוס, אך יש להדגיש בזהירות: התחום עדיין רחוק מהקליניקה הרפואית, שנמצאת הרבה שנים מאחור, ביחס למדע, גם יש לציין שהמחקרים קטנים יחסית, ולעיתים מתבצעים מסיבות מובנות בפריפריה רחוקה, לכאורה להימנע מהשפעת התורמים הגדולים. מחקר אקראי כפול־סמיות מ־2024 דיווח על הארכת טלומרים במדדים מסוימים לאחר שישה חודשי שימוש בתוסף מבוסס אסטרגלוס [מקור1, מקור2 מקור3].
לסיכום
סקירות עדכניות על ביולוגיית הטלומרים מדגישות בדיוק את המתח הזה: טלומרים קצרים ודיספונקציונליים דוחפים תהליך "בלאי" ביולוגי: סנסנס, פיברוזיס והזדקנות; אך הפעלה לא־מבוקרת של טלומראז היא חלק מארגז הכלים של ממאירויות רבות.
ובשפה פשוטה יותר:
- אסטרגלוס ונגזרותיו (למשל cycloastragenol / TA-65) [מקור].
- בהחלט קיימים מחקרי אדם קטנים המערערים על המחשבה השולטת בניהול מחלות אשר מצביעים על הארכת טלומרים או על עלייה בפעילות טלומראז, אך עדיין הם לא הובילו לפרס נובל.
- הפעלת טלומראז איננה פעולה ניטרלית ביולוגית, ולכן אין לראות בה "פתרון פשוט" לשחיקת טלומרים.
- קרני 🌞שמש
- הפחתת ROS ו־Nrf2 🥦
- שינה / אור יום / עומס ביולוגי
- תזונה שמפחיתה דלקת ושחיקה
- רסברטרול וצירי עמידות תאית
- אסטרגלוס / TA-65 / הפעלת טלומראז — ההבטחה והסיכון.
בפקעית הכליה, לחץ ורעלנים עלולים להפוך את הטלומרים לנקודת תורפה
בפקעית הכליה נפגשים עומס המודינמי, שטף מטבולי, ריכוזים גבוהים של מטבוליטים ורעלנים, ופעילות חמצונית מתמשכת — שילוב העלול להפוך את הטלומרים לנקודת תורפה ביולוגית. בניגוד לרושם הרווח, הפגיעה בפקעית אינה נגזרת רק מלחץ הדם הכללי הנמדד בזרוע, אלא גם, ולעיתים בעיקר, מן הלחץ השורר בתוך נימי הפקעית עצמה. לחץ זה — הלחץ התוך־גלומרולרי — נקבע לא רק על פי לחץ הדם הסיסטמי, אלא גם על פי טונוס העורקיק הנכנס והיוצא, היפרפילטרציה, עומס מטבולי (שפע תזוני [מקור]), מערכת רנין–אנגיוטנסין, רמות אינסולין, השמנה, סוכרת, ועומסים תזונתיים שונים, כמו יתר חלבונים, יתר סוכר ופחמימות. משום כך ייתכן בהחלט מצב שבו לחץ הדם הכללי אינו נראה קיצוני, אך בתוך יחידת הסינון עצמה שורר לחץ יתר כרוני, המעמיס על האנדותל, על הפודוציטים ועל המסנגיום, ומאלץ את הפקעית לפעול לאורך זמן בתנאים של מתיחה, חיכוך ועקה ביולוגית.
אחד הגורמים היכולים להחריף מצב זה הוא עומס חלבוני, ובייחוד חלבון מן החי, אשר זה יותר ממאה וחמישים שנה מתואר כגורם המגביר זרימת דם כלייתית והיפרפילטרציה [מקור1, מקור2, מקור3].
אין מדובר רק בשאלה של "כמה חלבון אוכלים", אלא באירוע פיזיולוגי רחב יותר: עלייה בזרימת הדם אל הפקעית, הגברת הלחץ על מחסום הסינון, והעמסת יתר על התאים האחראים לשמור על שלמותו. לכך מצטרפים גורמים נוספים — עודף חומצת שתן, מטבוליטים אורמיים, תוצרי גליקציה, עומס שומני, ולעיתים גם תוצרים פרו־דלקתיים שמקורם במזון ובמיקרוביום, כגון TMAO — וכל אלה יחד עלולים ליצור בפקעית סביבה שבה הלחץ המכנִי, העקה החמצונית והדלקת מזינים זה את זה.
במצב כזה, הנזק אינו מסתכם בפגיעה תפקודית זמנית או ב"עייפות" של הכליה, אלא עלול לחדור עד לרמת תחזוקת הגנום עצמה.
הלחץ התוך־גלומרולרי הגבוה והמתיחה המתמשכת של תאי האנדותל והפודוציטים מגבירים יצירת רדיקלים חופשיים, משבשים את ההומאוסטזיס המיטוכונדריאלי ומפעילים מסלולי דלקת.
במקביל, רעלנים ומטבוליטים המצטברים בדם וברקמה מחלישים את מערכות ההגנה של התא, פוגעים בנוגדי החמצון הטבעיים ומגדילים עוד יותר את שטף ה־ROS. כאן נכנסים הטלומרים אל מרכז הבמה: רצפים אלה, היושבים בקצות הכרומוזומים, עשירים בגואנין ולכן רגישים במיוחד לנזק חמצוני. חמצון של בסיסי גואנין, ובהם יצירת 8-oxoG, עלול להפריע לשכפול התקין של הטלומר, לערער את יציבותו, ולהחליש את ההגנה שמספק קומפלקס ה־shelterin — אותו מערך חלבונים האמור להסתיר את קצה הכרומוזום מפני מנגנוני תיקון העלולים לזהותו בטעות כשברדו־גדילי.
כאשר ההגנה הטלומרית נחלשת, קצה הכרומוזום עלול להיתפס כאתר נזק DNA. מכאן הדרך קצרה להפעלת תגובת נזק כרונית, לעצירת מחזור, לתהליך "בלאי" ביולוגי: סנסנס ולהפרשת אותות דלקתיים ופיברוטיים. במילים אחרות, הפקעית אינה נפגעת רק מפני ש"הלחץ גבוה" או מפני ש"רעלנים מסתובבים בדם", אלא מפני ששילוב של לחץ תוך־גלומרולרי, היפרפילטרציה, עקה חמצונית ועומס רעלי־מטבולי עלול לשחוק את מנגנוני ההגנה העדינים ביותר של התא — ובהם גם ההגנה על קצות הכרומוזומים עצמם. כאשר תהליך זה נמשך חודשים ושנים, התא הכלייתי עלול לחדול מהתחדשות, להיכנס למצב של הזדקנות תאית, ולהפוך ממבנה מסנן ומתפקד למקור של אותות המחמירים דלקת, צלקת ואובדן נפרונים.
המשמעות של כל זה איננה תיאורטית בלבד. הגורל הגנטי במקרה זה נדיר, והפך ככל הנראה סיסמת הסתרה, אבל אם הפגיעה בטלומרים בפקעית ניזונה לא רק מגורל גנטי, אלא גם מתנאים מקומיים של לחץ, היפרפילטרציה, רעלנים, עודף חלבון בלתי מותאם, חומצת שתן, מטבוליטים דלקתיים ופגיעה מיטוכונדריאלית — הרי שהדיון בטלומרים אינו יכול להסתיים בתיאור מנגנון השחיקה. הוא מחייב מעבר לשאלה המעשית: כיצד מפחיתים את תנאי השחיקה עצמם? כיצד מקטינים את הלחץ התוך־גלומרולרי ולא רק את לחץ הדם הכללי; כיצד מצמצמים היפרפילטרציה; כיצד מפחיתים את העומס המטבולי והרעלי המזין ROS; וכיצד מחזקים את מערכות ההגנה של התא לפני שקצות הכרומוזומים מאבדים את חסינותם. מכאן נפתח הצעד הבא: לא עוד טלומרים לבדם, אלא בחינת המערכות שאמורות לבלום את הנזק לפני שהוא מתקבע — גלוטתיון, NRF2🥦, שיקום מאזן החמצון, צמצום העקה המיטוכונדריאלית (Q10) והפחתת העומס הדלקתי והרעלי (שורש כורכום). רק בתוך מסגרת כזו ניתן להתחיל לדבר לא על נס, אלא על עבודה שיטתית שמטרתה לשמר בתא הכלייתי את היכולת לתקן, להתחדש ולהימנע מצלקת.
באילו תנאים חי התא הכלייתי עצמו. ובנושא זה האחריות עוברת אל מבקש השיקום.
תזכורת: המידע המוצג במאמר הוא בעל אופי מדעי ופילוסופי כללי, ואין לראות בו אבחון או המלצה לטיפול רפואי אישי. בכל מצב של מחלת כליות כרונית או פגיעה בתפקוד הכליתי, יש להיוועץ ברופא מומחה או בנפרולוג על מנת להתאים את הטיפול והתזונה למדדים האישיים של המטופל.
בכליה, ובייחוד בפקעית, הטלומרים אינם נשחקים בחלל ריק. הם מצויים בתוך רקמה הנתונה לעיתים ללחץ תוך־גלומרולרי גבוה, להיפרפילטרציה, לעודף חלבון בלתי מותאם, לרעלנים אורמיים, לחומצת שתן, לתוצרי גליקציה, ל־TMAO, לפגיעה מיטוכונדריאלית ולשטף ROS. לכן גם אם חומר כלשהו מסוגל לעורר טלומראז או להאט קיצור טלומרים בלויקוציטים, אין בכך עדיין הוכחה שהוא מגן על הפקעית או משקם תא אנדותל, פודוציט או תא אבוביתי החשופים מדי יום לעומס מכני, מטבולי וחמצוני. במילים אחרות, הארכת טלומרים בדם איננה שקולה לשיקום הטלומרים בכליה, ובוודאי לא להחלמה של רקמה שכבר מצויה במסלול של תהליך "בלאי" ביולוגי: סנסנס, דלקת ופיברוזיס. לשם כך דרושה מערכת שלמה של הפעלת צירי ההחלמה הביולוגיים שפועלים בגוף:
- צירי החלמה – מבוא למחשבה חדשה על בריאות והתחדשות מאת ירון מרגולין
- מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין.
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ב', על ההחלמה מאת ירון מרגולין
- חלק ג' – על ההחלמה
- חלק ד' – על שומר החומר הגנטי🔹🐦🔥 P53 🐦🔥 והנחיות טבעיות של הגוף ל: "החלמה".
- חלק ה' – חלבוני ההחלמה: הרשת המולקולרית של הריפוי, ההתחדשות וההומאוסטזיס מאת ירון מרגולין
- חלק ו' על צירי ההחלמה ומה שמפעיל אותם – (signal architecture) מאת ירון מרגולין
- 🔹 מפת מסלולי ההחלמה – 3 צירים – מבט כללי
- המסלולים הביולוגיים המשקמים את🔹 הגוף: פרספקטיבה חדשה על החלמה – מבט עומק על על 3 המסלולים המרכזיים.
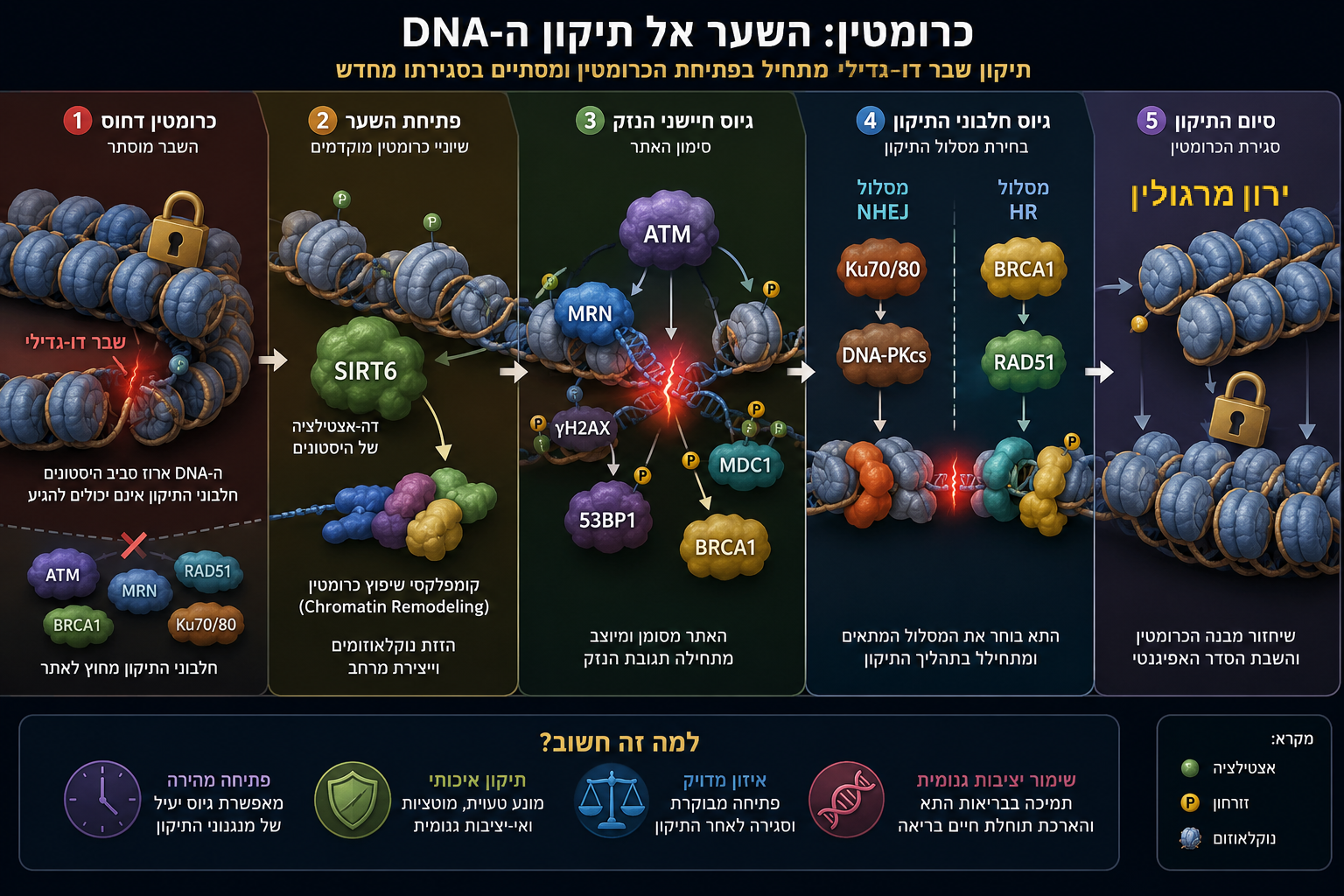
הדרך משבר דו־גדילי (DSB) ועד לשיקום היציבות הגנומית עוברת דרך פתיחת הכרומטין, גיוס מנגנוני התיקון 🛠️, ובנייה מחדש של המבנה האפיגנטי.
רק כאשר כל שלבי המסע מצליחים, התא יכול לבחור בהתחדשות במקום בסנסנס ובפיברוזיס.
כרומטין: השער אל התיקון
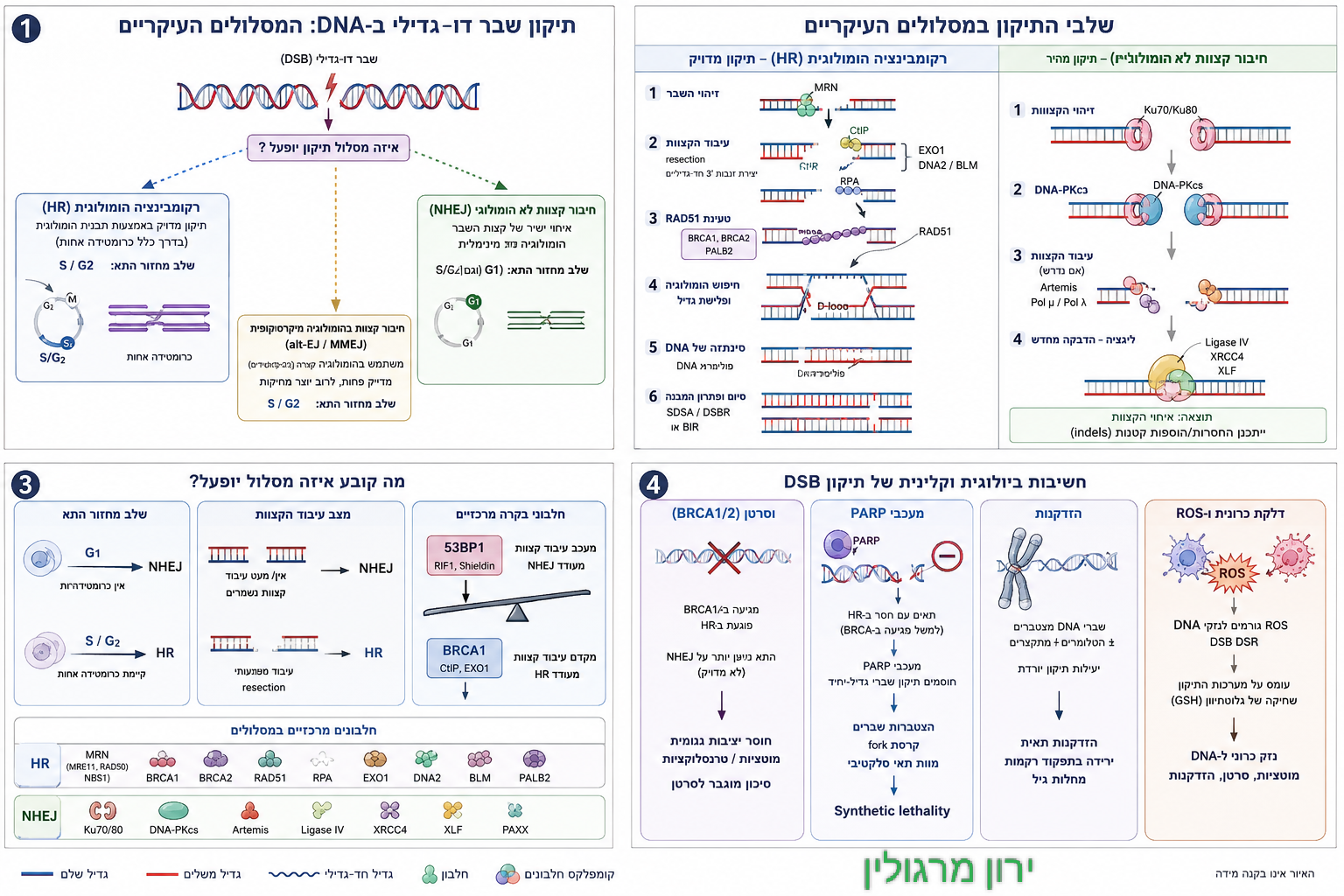
כאשר נוצר שבר דו־גדילי (Double-Strand Break, DSB), קל לדמיין את מנגנוני התיקון ממהרים אל מקום הפגיעה ומתחילים לחבר מחדש את ה־DNA. אולם במציאות, זהו אינו הצעד הראשון. לפני שחלבוני התיקון יכולים לזהות את השבר, לפני שהמערכת בוחרת בין HR ל־NHEJ, ולפני שמתחיל תהליך השחזור עצמו –
על התא לפתור בעיה בסיסית יותר: כיצד להגיע אל מקום הנזק.
ה־DNA האנושי אינו מונח בגרעין כחוט פתוח ונגיש. אורכו הכולל בכל תא מגיע לכשני מטרים, אך הוא ארוז בתוך גרעין שקוטרו מיקרונים ספורים בלבד. אריזה זו מתאפשרת באמצעות חלבוני היסטון, שסביבם נכרך ה־DNA ליצירת נוקלאוזומים, ואלה מאורגנים יחד למבנה מורכב המכונה כרומטין [מקור]. מבנה זה מעניק יציבות ומגן על המידע הגנטי, אך באותה מידה הוא גם מגביל את הגישה אליו.
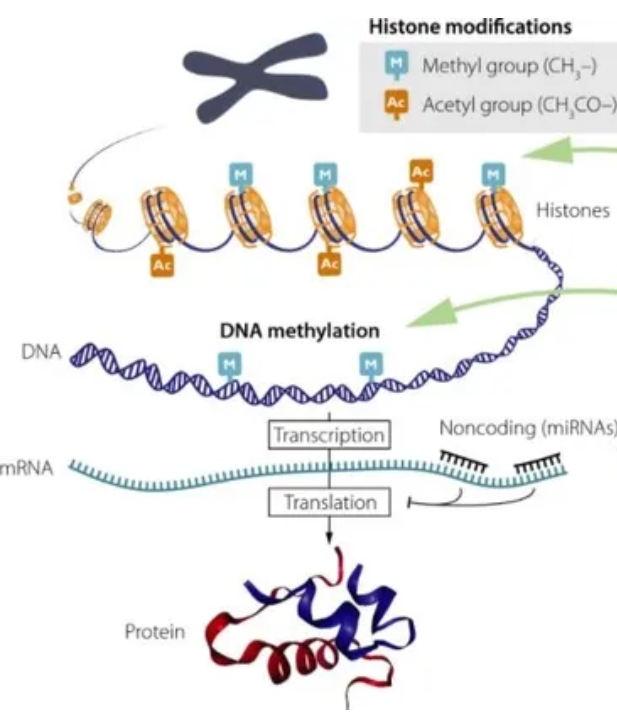
לכן, כאשר מתרחש שבר דו-גדילי, האתגר הראשון של התא אינו התיקון עצמו, אלא פתיחת הכרומטין באזור הפגיעה [מקור]. ללא פתיחה מבוקרת של המבנה הדחוס, חיישני הנזק וחלבוני התיקון 🛠️ אינם יכולים להגיע אל קצות ה־DNA השבורים, לזהות את אופי הפגיעה ולגייס את מנגנוני השיקום המתאימים.
שלב זה, המכונה Chromatin Remodeling, הוא אחד האירועים המוקדמים והחיוניים ביותר בתגובת נזק ה־DNA. שינויים כימיים בהיסטונים – כגון אצטילציה, דה־אצטילציה, מתילציה, זרחון ויוביקוויטינציה – משנים את מידת הדחיסות של הכרומטין ומאפשרים פתיחה זמנית ומבוקרת של אזור הנזק. במקביל, קומפלקסים ייעודיים לשיפוץ כרומטין מזיזים נוקלאוזומים ומפנים מקום למערכת התיקון.
רק לאחר שנפתחת "זירת האירוע" יכולים להיכנס לפעולה חלבוני החישה הראשונים. בין הראשונים שבהם נמצאים קומפלקס MRN והקינאז ATM, המזהים את השבר ומתחילים תגובת שרשרת מתואמת. בהמשך מגויסים חלבוני בקרה 🔍 כגון γH2AX, MDC1, 53BP1, BRCA1 וחלבונים נוספים, אשר מסמנים את אזור הפגיעה, מייצבים את התגובה ומכוונים את התא למסלול התיקון המתאים 🛠️.
במובן זה,
הכרומטין אינו רק הרקע שעליו מתרחש התיקון – הוא השער ⛩️ שדרכו כל תהליך התיקון מתחיל.
כאן מתברר גם תפקידו המרכזי של SIRT6 🫐. מחקרים מצביעים על כך שחלבון זה מגויס במהירות לאתרי שבר דו־גדיליים (DSB), שם הוא משנה את מבנה הכרומטין באמצעות דה־אצטילציה של היסטונים, תומך בגיוס קומפלקסי שיפוץ כרומטין, ומאפשר לחלבוני התיקון 🛠️ להגיע אל אתר הנזק. במילים אחרות,
SIRT6 🫐 אינו "מדביק" את ה־DNA, אלא מארגן את זירת התיקון. ללא פעילותו, הגישה אל השבר עלולה להיות איטית, חלקית או בלתי מספקת, והתוצאה עלולה להיות תיקון שגוי, הצטברות נזק ואי־יציבות גנומית.
חשיבותו של הכרומטין אינה מסתיימת עם פתיחת האזור הפגוע.
לאחר השלמת התיקון,
על התא להשיב את הכרומטין למבנהו התקין.
כרומטין שיישאר פתוח לאורך זמן עלול לחשוף גנים שאינם אמורים להיות פעילים, לשבש את דפוסי הביטוי הגנטי ולהגביר אי־יציבות כרומוזומלית.
לכן, תיקון DNA איכותי אינו מסתיים באיחוי השבר, אלא 🏹 רק כאשר גם הארגון המרחבי והאפיגנטי של הכרומטין משוחזר במלואו.
נקודה זו חשובה. היא משנה את האופן שבו אנו תופסים את מנגנוני התיקון 🛠️.
במשך שנים רבות נתפסו חלבוני HR ו־NHEJ כ"גיבורי הסיפור". כיום מתברר ש
הם פועלים רק לאחר שהכרומטין עצמו מאפשר להם גישה.
למעשה, איכות התיקון תלויה לא רק באנזימי התיקון 🛠️, אלא גם במידת היכולת של התא לפתוח את הכרומטין בזמן ⛩️, לגייס את החלבונים הנכונים, ולאחר מכן לסגור מחדש את המבנה באופן שישיב את היציבות הגנומית⛔.
מכאן גם נובעת חשיבותה של הסביבה התאית. עקה חמצונית מתמשכת, פגיעה מיטוכונדריאלית, דלקת כרונית, ירידה בזמינות NAD⁺, ושינויים אפיגנטיים הנלווים להזדקנות עלולים לפגוע לא רק בחלבוני התיקון 🛠️ עצמם, אלא גם ביכולת לבצע את שלב פתיחת הכרומטין. כאשר "שער התיקון" נפתח באיחור ⛩️, באופן חלקי או שאינו נסגר כראוי, גם מנגנוני התיקון 🛠️ המתוחכמים ביותר אינם יכולים לפעול במלוא יעילותם ⛔.
הבנה זו מובילה אל השלב הבא בסיפור ההחלמה הטבעי. אם נגישות הכרומטין היא השער ⛩️ לתיקון 🛠️, מתעוררת השאלה מה קובע את מצבו של שער זה ⛩️ לאורך החיים. התשובה לשאלה זו מצויה במרחב המופלא של האפיגנטיקה – בדפוסי המתילציה, בסימוני ההיסטונים ובמערכת המורכבת המארגנת את הכרומטין בכל רגע נתון.
ג. SIRT1 ו־SIRT6: שומרי הכרומטין, מארגני תגובת התיקון, ומכריעי גורל התא
אם הטלומרים מציבים את גבול היכולת של התא להמשיך להתחלק מבלי לאבד את זהותו, הרי ש־SIRT1🍇 ו־SIRT6 🫐 מסייעים לקבוע מה יעשה התא כאשר נזק כבר הופיע: האם יגייס מערכות תיקון, יכבה דלקת, ייצב את הכרומטין וישוב למסלול של הישרדות — או שייכשל, יישאב לעקה כרונית, ויידחף לעבר סנסנס, פיברוזיס ואובדן תפקוד. שתי הסירטואינים הללו, חלבונים תלויי NAD⁺, אינן “עוד שחקן” ברשימת מנגנוני תיקון; הן מצויות קרוב מאוד לליבת ההכרעה של התא, משום שהן מחברות בין מצב האנרגיה, מצב הכרומטין, תגובת נזק ה־DNA, עקה חמצונית והזדקנות תאית.
כיצד נשמרת בכלל סביבת כרומטין שמאפשרת לזהות נכון את הנזק, לפתוח את האזור הדרוש לתיקון, ולגייס את המכונה המולקולרית המתאימה בזמן הנכון.
SIRT1 🍇 פועל במידה רבה כווסת של תוכנית ההישרדות התאית. הוא משפיע על שעתוק, על תגובות דלקת, על מסלולי עקה חמצונית ועל גורמי מפתח כמו p53🐦🔥, FOXO ו־NF-κB. דרך דה־אצטילציה של חלבונים אלה ואחרים, SIRT1 🍇 עשוי להחליש דחיפה מוקדמת מדי אל תהליך "בלאי" ביולוגי: סנסנס או אפופטוזיס, לחזק ביטוי של גנים נוגדי חמצון, ולתמוך במצב שבו התא עדיין “מאמין” שיש טעם לתקן. במקביל, SIRT6 פועל ישירות על הכרומטין ועל אתרי נזק: הוא משתתף בארגון מבנה הכרומטין, מסייע בגיוס חלבוני תיקון 🛠️, ומקושר הן למסלולי HR והן למסלולי NHEJ. במילים אחרות, אם פרק הטלומרים עסק בהגנה על קצות הכרומוזומים מפני זיהוי שגוי כנזק, הרי שכאן אנו עוברים לשאלה הרחבה יותר:
SIRT6 🫐: מן הכרומטין אל אתר השבר
SIRT6 🫐 (סירטואין 6) הוא חלבון הפועל בגרעין התא, ומתפקד כ"מפקח" או "וסת" מרכזי של תהליכים ביולוגיים רבים. הוא זוכה למחקר רב בשנים האחרונות בשל תפקידו הקריטי בשמירה על בריאות הגוף ואריכות ימים [מקור].
SIRT6 🫐 נחשב כיום לאחד החלבונים החשובים ביותר בקשר שבין יציבות כרומטין לתיקון שברים דו־גדיליים [מקור]. הוא פועל על היסטונים, ובכך משפיע על מידת הדחיסות של הכרומטין ועל נגישות ה־DNA למערכות התיקון. כרומטין “סגור” מדי עלול להקשות על גישה של חיישני הנזק ושל חלבוני התיקון 🛠️ אל אתר השבר; כרומטין כאוטי מדי, לעומת זאת, עלול לפגוע בדיוק הזיהוי ולהעצים אי־יציבות גנומית. SIRT6 🫐 משתתף באיזון העדין הזה: הוא מסייע לעצב כרומטין שמצד אחד שומר על יציבות, ומצד אחר מסוגל להיפתח במהירות כאשר מופיע שבר.
ברגע שנוצר DSB, התא אינו זקוק רק ל”חומרי תיקון”, אלא גם לארגון מרחבי נכון של האתר הפגוע [מקור]. כאן נכנס SIRT6 🫐 לתמונה. מחקרים מצביעים על כך שהוא מגויס לאתרי שבר, תומך בהפעלת תגובת ATM, ומסייע בגיוסם או בפעילותם של חלבוני תיקון מרכזיים. במצבים מסוימים הוא מקושר לחלבוני NHEJ כגון DNA-PKcs, ובמצבים אחרים לפקטורים של HR, כך שהוא אינו “חלבון של מסלול אחד”, אלא יותר מארגן־על של סביבה מתקנת. משמעות הדבר עמוקה: הבחירה בין HR ל־NHEJ אינה נקבעת רק על ידי שלב מחזור התא, אלא גם על ידי השאלה אם התא הצליח לבנות סביב השבר כרומטין נגיש, מסומן כראוי, עתיר חלבוני תיקון 🛠️ ובעל אספקת אנרגיה מספקת🔋.
כאשר SIRT6 🫐 חסר, נפגע או מדוכא, מתרבים סימני נזק DNA, עולה אי־היציבות הגנומית, והתא נוטה יותר להזדקנות מוקדמת. לכן אין זה מפתיע ש־SIRT6 🫐 נקשר שוב ושוב גם להזדקנות מערכתית וגם לפגיעה באיברים עתירי עומס מטבולי. בכליה, משמעות הדבר עשויה להיות חריפה במיוחד: תא כלייתי שנחשף שוב ושוב לעקה חמצונית, לרעלנים אורמיים, לעומס מטבולי ולפגיעה מיטוכונדריאלית, זקוק לא רק ליכולת “לתקן שבר”, אלא ליכולת לשמר ארכיטקטורת כרומטין מתקנת לאורך זמן. ללא יכולת זו, גם שברים קטנים יחסית עלולים להפוך לאות כרוני של סכנה, לדחוף את התא לסנסנס ופיברוזיס.
SIRT1🍇: לאפשר לתא להישאר בר־תיקון
אם SIRT6 🫐 פועל קרוב מאוד לאתר השבר ולמבנה הכרומטין, SIRT1 🍇 פועל ברמה רחבה יותר של “החלטת התא” אם להישאר במסלול בר־תיקון. הוא אינו מתקן שבר בידיו, אלא מסייע לעצב את מצב התא שבתוכו תיקון עדיין אפשרי. SIRT1 🍇 מדכא בחלק מן ההקשרים פעילות עודפת של p53, מווסת את FOXO, מפחית איתות דלקתי דרך NF-κB, ותומך בביטוי של מערכות נוגדות חמצון ובהסתגלות לעקה מטבולית. במצב כזה, התא עשוי להישאר פחות דלקתי, פחות מחומצן, ויותר פנוי להשקיע משאבים בתיקון ולא בכניסה למצב חירום מתמשך.
במילים אחרות, SIRT1 🍇 אינו רק “חלבון אנטי־אייג’ינג”, אלא חלק ממנגנון שמגדיר אם סביבת התא תהיה סביבת תיקון או סביבת קריסה. כאשר רמות ROS גבוהות, המיטוכונדריה פגועות, מאגרי NAD⁺ נשחקים והדלקת הכרונית פעילה, SIRT1 🍇 נחלש — ואז התא מאבד חלק מיכולתו לבלום את מעגל ההסלמה. p53 🐦🔥 עלול להישאר פעיל לאורך זמן, אותות דלקתיים מתגברים, ותגובות הסתגלותיות כמו FOXO וניקוי נזק חמצוני פועלות בעוצמה נמוכה יותר. התוצאה עלולה להיות מעבר הדרגתי ממצב של תיקון למצב של סנסנס, שבו התא חדל להתחלק, מפריש ציטוקינים פרו־דלקתיים, ומעמיק את הפגיעה ברקמה שסביבו.
בכליה, זו אינה שאלה תאורטית. פודוציטים, תאי האבובית ותאי האנדותל נאלצים להתמודד עם עומס חמצוני, תנודות מטבוליות, דלקת, לעיתים היפוקסיה מקומית ולעיתים גם חשיפה לרעלנים אורמיים. בסביבה כזו, חלבון כמו SIRT1 עשוי להיות ההבדל בין תא שמצליח לבלום הסלמה ולשמר גמישות מטבולית — לבין תא שנלכד במעגל של ROS, דלקת, נזק DNA וסנסנס. מכאן שהשאלה החשובה איננה רק “האם יש שבר דו־גדילי”, אלא באיזה מצב פוגש התא את השבר: האם הוא מצויד ב־NAD⁺, במיטוכונדריות מתפקדות, בגלוטתיון ובמערכת נוגדת חמצון פעילה — או שהוא כבר מותש, מחומצן ודלקתי.
- מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין.
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ב', על הפעלתם של צירי ריפוי טבעי בגוף ועל אפשרות להגיע להחלמה מאת ירון מרגולין
- חלק ג' – על הפעלת מהלכי ההחלמה
- חלק ד' – על שומר החומר הגנטי🔹🐦🔥 P53 🐦🔥 והנחיות טבעיות של הגוף ל"החלמה".
- חלק ה' – חלבוני ההחלמה: הרשת המולקולרית של הריפוי, ההתחדשות וההומאוסטזיס מאת ירון מרגולין
- חלק ו' על צירי ההחלמה ומה שמפעיל אותם – (signal architecture) מאת ירון מרגולין
- מפת מסלולי ההחלמה – 3 הצירים המרכזיים – מבוא לצירי ההחלמה.
- המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה – מבט עומק על על 3 המסלולים המרכזיים.
SIRT1🍇, SIRT6🫐 ותגובת ATM: לא רק זיהוי נזק, אלא ארגון של תגובה שלמה
🚨 תגובת ATM: האזעקה המולקולרית המרכזית של הגנום
חלבון ה-ATM (Ataxia Telangiectasia Mutated) הוא הקינאז המרכזי והמפקד המולקולרי בשטח ברגע שבו מתרחש שבר דו-גדילי (DSB) ב-DNA. במצב שגרה,
קיים בתא בצורה פסיבית כצמד חלבונים קשורים (דימר). אולם, ברגע שחלה קטיעה פיזית של שני גדילי ה-DNA, קומפלקס חלבונים ייעודי מזהה את הקצוות החשופים ומגייס את ATM אל נקודת השבר. מפגש זה גורם ל-ATM לעבור שינוי מבני מיידי, להתפרק לחלבונים בודדים (מונומרים) ולעבור זרחון עצמי שמפעיל אותו לחלוטין.
ברגע שנכנס לפעולה, ATM מתפקד כ"מערכת כריזה" תאית רבת עוצמה. הוא מזרחן מאות חלבוני מטרה בתוך שניות, ובראשם את חלבון המבנה H2AX (שהופך ל-γH2AX), ובכך הוא מסמן ומאיר את מיקום האסון עבור אנזימי התיקון של ה-HR וה-NHEJ. במקביל, ATM מזרחן ומייצב את חלבון ה-p53🐦🔥, מה שמפעיל את ה"ברקס" התאי p21 ועוצר את מחזור התא כדי למנוע חלוקה הרסנית.
הזווית הכלייתית: כדי ש-ATM יוכל לנווט במהירות אל נקודת השבר, מבנה הכרומטין (האופן שבו ה-DNA ארוז) חייב להיות גמיש ונגיש. פגיעה במסלולי המתילציה אצל חולי כליות משבשת את הארכיטקטורה האפיגנטית של הכרומטין, מה שעלול למנוע מ-ATM להגיע לשבר או להפעיל את איתות האזעקה בצורה יעילה. כתוצאה מכך, שברים דו–גדיליים (DSB) נותרים פתוחים, הם אינם מתוקנים, ובהמשך הם מדרדרים את תאי הכליה במהירות אל עבר מסלול הסנסנציה (הזדקנות תאית) והפיברוזיס ההרסני. רפואת הממסד מתבוננת הלומה נוכח התופעה וזועקת בכל כוחה, אבל אין שומע.
תגובת ATM היא אחת מנקודות המפתח במפגש של התא עם שבר דו–גדילי (DSB). כפי שרשמתי למעלה היא מפעילה שרשרת אירועים שמסמנת את אזור הנזק, מעכבת את מחזור התא, ומאפשרת גיוס של מנגנוני תיקון. אך ATM אינו פועל בריק. כדי שתגובתו תהיה יעילה, יש צורך בכרומטין 🎁 שמסוגל להיפתח, בסביבה ביוכימית שאינה מוצפת מדי ב־ROS, ובמערכת בקרה שאינה דוחפת את התא מהר מדי אל תהליך "בלאי" ביולוגי – סנסנס. כאן בדיוק מצטלבים תפקידי SIRT1🍇 ו־SIRT6🫐: האחד תומך בסביבת הישרדות ותיקון רחבה, והשני תומך בארגון המקומי של הכרומטין ובגיוס מנגנוני תיקון אל אתר השבר.
מכאן עולה נקודה עקרונית למאמר כולו: תיקון DNA אינו רק מנגנון אנזימטי, אלא מצב מערכתי של התא. גם אם קיימים חלבוני HR ו־NHEJ, אין די בכך. התא צריך כרומטין מאורגן, מאגרי NAD⁺ מספקים, אותות דלקת מאוזנים, מיטוכונדריות שאינן קורסות תחת עומס ROS, והפעלה של מסלולי הישרדות שאינם דוחפים אותו במהירות אל זקנה תאית. SIRT1🍇 ו־SIRT6 🫐 ממחישים זאת היטב: הם אינם “תוספת” למסלולי התיקון, אלא חלק מהקרקע שעליה בכלל יכולה לצמוח תגובת תיקון אפקטיבית.
מן הסירטואינים אל הקרקע האפיגנטית של התיקון
אלא שגם SIRT1🍇 ו־SIRT6🫐 אינם פועלים בחלל ריק. פעילותם קשורה קשר עמוק למצב הכרומטין, לסימוני היסטונים, לדפוסי מתילציה ולשאלה אילו אזורים בגנום פתוחים, סגורים או מסומנים כראוי לתגובה. במילים אחרות, לאחר שהבנו כי טלומרים מגינים על קצות הכרומוזומים, וכי הסירטואינים מסייעים לארגן את תגובת התא לנזק ולשמר אותו במצב בר־תיקון — עלינו לעבור אל השכבה הבאה: הקרקע האפיגנטית שעליה פועלים מנגנוני התיקון עצמם.
להלן כמה מן המולקולות הטבעיות שנחקרו כמודולטורים של SIRT6, בעיקר במערכות ניסוי תאיות וביוכימיות – מקדמי הפעלת SIRT6:
הציאנידין
מוכרים לחוקרים כיום כבר מספר חומרים טבעיים ופשוטים שהוכחו על ידם כמפעילי SIRT6🫐, כאשר המקבילה הבולטת ביותר לרסברטרול🍇 שמוכר כמפעיל של SIRT1🍇 הוא הציאנידין (Cyanidin) ומספר פלבנואידים נפוצים אחרים [מקור1, מקור2].
1. ציאנידין (Cyanidin) – המפעיל העוצמתי ביותר
- ציאנידין הוא פיגמנט צמחי טבעי (אנתוציאנין) המעניק את הצבע האדום-סגול הכהה לפירות.
- הקשר ל-SIRT6 🫐: במחקרים נמצא כי ציאנידין הוא מפעיל עוצמתי במיוחד, אשר הגביר את פעילות SIRT6 פי 55 (לשם השוואה, חומרים אחרים הגבירו אותה פי 3 עד 10 בלבד).
- מקורות טבעיים: פירות יער (אוכמניות🫐, פטל שחור), ענבים כהים, חמוציות, רימונים ותפוחים אדומים🍎. [מקור1, מקור2].
קוורצטין, איזוקוורצטין ו־לוטאולין — פלבנואידים המצויים, בין השאר, בבצל אדום, צלפים, תפוחים, כוסמת ירוקה, סלרי, פטרוזיליה (אסורה לחולי כליה) ופלפלים — ונמצא כי הם עשויים להיקשר ל־SIRT6 ולשנות את פעילותו לטובה.
2. קוורצטין 🧅(Quercetin) ואיזוקוורצטין
- הקוורצטין 🧅 הוא נוגד חמצון פלבנואידי נפוץ ומוכר מאוד בעולם הפעלת נתיבי ההחלמה בעזרת התזונה כתרופה.
- הקשר ל-SIRT6: מחקרים מבניים הראו כי קוורצטין 🧅 , תלוי ריכוז, מסוגל להיקשר ישירות ל-SIRT6 🫐 ולהפעיל אותו. הגרסה הטבעית שלו, איזוקוורצטין (Isoquercetin), נחשבת למפעילה ספציפית ומדויקת עוד יותר של חלבון זה.
- מקורות טבעיים: בצל אדום, צלפים, תפוחים🍎, כוסמת, ועלים ירוקים🥬 [מקור1, מקור2, מקור3]. קמח קליפות בצל, תה מקליפות בצל [מקור]
3. לוטאולין 🫑 (Luteolin) ופיזטין (Fisetin) 🍓
- לוטאולין ופיזטין אלו הם פלבנואידים צמחיים המוכרים בזכות תכונותיהם האנטי-דלקתיות ומפחיתות סטרס חמצוני.
- הקשר ל-SIRT6: שניהם הראו יכולת להגביר את הביטוי של SIRT6, ובכך לתמוך במנגנוני הגנה על התא.
- מקורות טבעיים: סלרי, פטרוזיליה, פלפל ירוק 🫑 (עבור לוטאולין), ותות שדה (עבור פיזטין) 🍓 [מקור1, מקור2].
- נמצא כי פיזטין פלבנואיד טבעי המצוי בעיקר בתותים, תפוחים🍎, ענבים, בצל ומלפפונים🥒 בעל חשיבות רחבה יותר בתהליך "בלאי" ביולוגי: סנסנס, עקה חמצונית ודלקת, עם אפשרות להשפעה עקיפה או הקשרית על מסלולי סירטואינים — למרות שפיזטין מוכר בעיקר כתרכובת נוגדת סננסנס, עקה חמצונית ודלקת, יש גם עדות ישירה ומעניינת לקשר שלו ל-SIRT6. במחקר עדכני [מקור] נמצא שפיזטין העלה את פעילות/ביטוי SIRT6 בתאי סחוס, וחלק מההשפעות המגנות שלו נחלשו כאשר SIRT6 עוכב, כך שניתן לטעון על השפעה אם כי המחקר מצביע — על הקשר משני ורקמתי יותר מאשר מיצוב חד-משמעי של פיזטין כ”מפעיל SIRT6 קלאסי [מקור].
פוקוידאן:
מודולטור נוסף שנחקר בהקשר של SIRT6 הוא פוקוידאן מאצות חומות, אך משמעותו הביולוגית והקלינית עדיין רחוקה מהכרעה [מקור1, מקור2].
4. פוקוידאן (Fucoidan)
- פוקוידאן -פחמימה מורכבת (רב-סוכר) המצוי באצות ים.
- הקשר ל-SIRT6: מחקרים עדכניים מהשנים האחרונות זיהו את הפוקוידאן כמפעיל ישיר של SIRT6 [מקור1, מקור2]. בניסויים בבעלי חיים, הפעלת החלבון באמצעות פוקוידאן הובילה להארכת תוחלת החיים הבריאה ולשיפור מדדים מטבוליים.
- מקורות טבעיים: אצות חומות (כמו קומבו, וואקאמה וחיג'יקי). [מקור]
5. רסברטרול 🍇 עצמו (Resveratrol)
- SIRT1🍇 מוכר בעיקר כמפעיל של SIRT1🍇, מחקרים מראים כי במינונים מסוימים הוא מסוגל להיקשר ולהפעיל גם את SIRT6, אם כי לא בבאופן בלעדי. [מקור1, מקור2].
ד. מתילציה, כרומטין ויציבות גנומית: הקרקע שעליה פועלים מנגנוני התיקון
אם SIRT1 ו־SIRT6 מייצגים שכבת בקרה דינמית שמגיבה לנזק DNA ומארגנת את סביבת התיקון בזמן אמת, הרי ש־המתילציה של ה־DNA וההיסטונים, יחד עם ארגון הכרומטין, מגדירים את התנאים הקבועים יותר שבהם בכלל מתרחשת תגובת הנזק שמובליה גם לשבר דו–גדילי (DSB). במילים אחרות, לא רק השאלה כיצד התא מתקן שבר קובעת את גורלו, אלא גם השאלה:
באיזה נוף אפיגנטי השבר הזה מתרחש.
- אפיגנטיקה ומזון כתרופה ביולוגית להחלמה ממחלות גנטיות
- באיזה אופן תהליך המתילציה משפיע על ההחלמה מחלות כרוניות
- אפיגנטיקה ומזון כתרופה ביולוגית להחלמה ממחלות גנטיות
- שמונה עקרונות ההחלמה
- המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה
- עורקים גמישים – הסוד והדרך לזכייה בבריאות מחדש
כרומטין אינו רק “אריזה” של DNA, אלא מערכת רגולטורית שמחליטה אילו אזורים יהיו נגישים לשעתוק, אילו גנים יישארו שקטים, ואילו אזורים יהיו זמינים לגיוס מהיר של מערכות תיקון. כאשר דפוסי היסטונים נשמרים בצורה מאורגנת, התא מסוגל להבחין בין אזורים פעילים לאזורים דחוסים, ולכוון את תגובת ה־DNA damage response באופן מדויק. אך כאשר המערכת האפיגנטית מתערערת, הגבולות הללו מיטשטשים:
אזורי כרומטין “נפתחים” באופן לא תקין או נסגרים באופן פתולוגי, ותפקוד מנגנוני התיקון הופך פחות מדויק.
במצב תקין, דפוסי מתילציה פועלים כמעין “זיכרון יציב” של זהות תאית. הם שומרים על כך שתאי פודוציט שבכליה יישארו פודוציטים, שתאי אבובית שבכליה, ישמרו על פרופיל מטבולי מתאים, ושמערכות תגובת עקה לא יופעלו ביתר. אך עם הזמן, מתקדמת הצטברות רעלים ממקור מטבולי – תחת עומס של ROS, פגיעה מיטוכונדריאלית, דלקת כרונית ולחץ מטבולי מתמשך, מה שמכונה באופן כללי תופעת הגיל… מתרחשת התרופפות הדרגתית של הסדר האפיגנטי. תהליך זה אינו רק שינוי ביטוי גנים, אלא שינוי עומק במבנה שבו ה־DNA עצמו “מוצג” למערכות התיקון. בשלב זה מערך ההנחיות (החומר הגנטי) משתבש ומתחילה תשדורת פעולה שגויה שהיא עלולה פהיות הרסנית.
כאשר הכרומטין מאבד את יציבותו, גם תגובת שבר דו–גדילי (DSB) נפגעת. חלבוני חישה מתקשים לזהות הקשרים נכונים, גיוס של ATM וגורמי תיקון הופך פחות יעיל, והבחירה בין HR ל־NHEJ עלולה להיות פחות מדויקת. במקום תיקון נאמן לתבנית, התא נוטה יותר למסלולים מהירים עליהם העמקנו בחלק א' של המאמר [מקור] אך שגויים, מה שמגדיל את הסיכון לשגיאות תיקון, שברים כרומוזומליים משניים ואי־יציבות גנומית מצטברת.
כאן יש להדגיש כי:
האפיגנטיקה אינה רק “רקע” לתיקון DNA — והחזרת הוראות ההפעלה הנכונות לגוף, היא חלק ממנגנון ההחלטה עצמו.
כאשר אזורי כרומטין פתוחים מדי, מתקבלת לעיתים תגובת יתר לנזק, עם הפעלה לא מבוקרת של מסלולי עקה. כאשר הם סגורים מדי, נזק אמיתי עלול “להיעלם” מהמערכת ולהתגלות מאוחר מדי, כאשר כבר הצטבר לכדי קריסה פונקציונלית. שני המצבים הללו מובילים לאותה תוצאה סופית: אובדן איזון בין תיקון, הישרדות והתחדשות.
בכליה, הרגישות לתהליך זה גבוהה במיוחד. תאי הכליה פועלים בסביבה של תנודות חמצן, עומס מטבולי גבוה וחשיפה מתמשכת למולקולות שעלולות לפגוע ב־DNA.
כאשר דפוסי המתילציה יציבים, התא מסוגל לשמור על תגובת תיקון מדויקת גם תחת עומס.
אך כאשר הסדר האפיגנטי נשחק, כל אירוע של נזק DNA עלול להטות את התא מעט יותר לכיוון של תגובת יתר, דלקת כרונית או כניסה למסלול של יצירת "בלאי" ביולוגי – סנסנס.
מכאן עולה קשר ישיר בין אפיגנטיקה לבין תפקוד מערכות התיקון עצמן:
לא ניתן להבין HR, NHEJ או תגובת ATM במנותק מהשאלה כיצד נראה הכרומטין שבו הם פועלים. ה־DNA אינו “נחשף” לתיקון במצב ניטרלי — הוא תמיד מופיע בתוך הקשר אפיגנטי שמכתיב את איכות התגובה.
בנקודה זו מתחדד הקשר בין שכבות המאמר כולו: אם הטלומרים מגדירים את גבולות הזמן של התא, והסירטואינים (🍇🫐) מווסתים את תגובת התיקון בזמן אמת, הרי שהאפיגנטיקה מגדירה את המרחב שבו כל זה מתרחש. ללא מרחב יציב כזה, גם מערכות תיקון מתקדמות 🛠️ אינן מבטיחות יציבות גנומית לאורך זמן.
מכאן טבעי ונכון להדגיש שגם מצב חמצוני ואנרגטי בהחלט מכתיב את עצם האפשרות לבצע תיקון:
ה- גלוטתיון, Nrf2🥦 והפחתת ROS: תנאי הסביבה של תיקון DNA 🧬
אם הכרומטין והאפיגנטיקה מגדירים את המרחב שבו מתרחש תיקון DNA, הרי ש
מערכת הגלוטתיון, Nrf2 🥦והוויסות של ROS מגדירה את איכות הסביבה שבה המרחב הזה מתקיים.
במילים אחרות: גם התא המאורגן ביותר אפיגנטית לא יוכל לתקן שברים דו־גדיליים (DSB) אם הוא מצוי בסביבה חמצונית כרונית שאינה מאפשרת יציבות מולקולרית.
ROS (Reactive Oxygen Species) אינם רק תוצר לוואי של מטבוליזם, אלא גורם ישיר לנזק DNA, כולל יצירת בסיסים מחומצנים כמו 8-oxoG, אשר מערערים את יציבות המידע הגנטי ומגדילים את הסיכון לשברים דו־גדיליים.
כאשר עומס ROS גבוה ומתמשך, הנזק אינו עוד אירוע נקודתי אלא מצב רקע כרוני. במצב כזה, מערכות התיקון 🛠️ אינן “מתקנות שבר”, אלא מנסות להתמודד עם שטף בלתי פוסק של פגיעות 🌪️.
כאן נכנסת מערכת Nrf2🥦 כציר מרכזי של תגובת ההגנה החמצונית.
Nrf2 🥦 מווסת את הביטוי של אנזימים נוגדי חמצון, כולל מערכות התומכות בסינתזה ובמחזור של גלוטתיון, שהוא אחד המגנים המרכזיים של התא מפני עקה חמצונית. כאשר Nrf2 🥦 פעיל באופן תקין, התא מסוגל להפחית את עומס ה־ROS, לייצב את סביבת המיטוכונדריה, וליצור תנאים שבהם נזק DNA נשאר ברמה ניתנת לניהול.

לעומת זאת, כאשר Nrf2 🥦 מדוכא או מותש — כפי שקורה לעיתים בתהליכי בלי מתקדמים מה שמכנים במחקר גם הזדקנות, דלקת כרונית או עומס מטבולי ממושך — נוצר מצב שבו המיטוכונדריה עצמה הופכת למקור עיקרי של נזק. אז מתחיל מעגל סגור: ROS פוגע ב־DNA, פגיעה ב־DNA מפעילה תגובות עקה, ותגובות העקה מחלישות עוד יותר את המיטוכונדריה ואת מערכות נוגדות החמצון.
במצב זה, גם מסלולי HR ו־NHEJ פועלים בתנאים קשים יותר. חלבוני תיקון נחשפים לנזק חמצוני, האיתותים של ATM ו־PARP הופכים כרוניים, ולעיתים התא נוטה יותר להפעלה מתמשכת של תגובות חירום במקום תיקון מדויק. התוצאה אינה רק הצטברות שברים, אלא שינוי מהותי במאזן בין תיקון, הישרדות והתפתחות "בלאי" ביולוגי – סנסנס.

בכליה, שבה קצב המטבוליזם גבוה במיוחד והחשיפה לעומסים חמצוניים ותזונתיים משמעותית, מערכת Nrf2 🥦–גלוטתיון אינה רק מערכת הגנה, אלא תנאי יסוד לקיום יציב של תאי רקמה לאורך זמן. כאשר מערכת זו נחלשת, כל שבר DNA הופך לאירוע בעל פוטנציאל מערכתי — ולא רק מקומי.
מכאן מתבקש המעבר לשכבה האחרונה של המסגרת המטבולית־אנרגטית של תיקון DNA:
ו. NAD⁺, PARP ומיטוכונדריה: האנרגיה של התיקון והמחיר של קריסה מטבולית
תיקון DNA, ובפרט תיקון שברים דו־גדיליים (DSB), הוא תהליך עתיר אנרגיה. הוא דורש לא רק חלבונים מתאימים, אלא גם זמינות גבוהה של NAD⁺, תפקוד מיטוכונדריאלי תקין, ואיזון עדין בין יצירת אנרגיה לבין תגובת עקה. במרכז צומת זה ניצב האנזים PARP, אשר מזהה נזק DNA ומפעיל שרשרת תגובות שמסייעות בגיוסם של מנגנוני תיקון 🛠️.
שבר דו–גדילי (DSB)
כאשר הנזק מתון ומוגבל, מערכת PARP פועלת באופן יעיל ותומכת בתיקון. אך כאשר נזק ה־DNA הופך כרוני, PARP עלול להפוך לצרכן כבד של NAD⁺. מצב זה יוצר ירידה במאגרי האנרגיה התאית, ומוביל לפגיעה בתהליכים תלויי NAD⁺ — כולל פעילות סירטואינים (🫐🫐🍇), תפקוד מיטוכונדריאלי🔋 ויכולת תגובה לעקה. במילים אחרות, מערכת שנועדה לתקן DNA עלולה, כאשר היא מופעלת יתר על המידה, להפוך לגורם שמחליש את יכולת התיקון עצמה.
המיטוכונדריה 🔋נמצאת במרכז המעגל הזה. היא לא רק ספקית אנרגיה, אלא גם רגולטורית של ROS ושל אותות מטבוליים המשפיעים על תיקון DNA. כאשר המיטוכונדריה 🔋פגועה, עולה ייצור ROS, יורדת זמינות ATP, ומצטמצמת היכולת לתמוך בתהליכי HR ו־NHEJ. במצב כזה, התא נכנס למעגל של עקה מטבולית: יותר נזק, פחות אנרגיה לתיקון, וירידה הדרגתית ביכולת לשמור על יציבות גנומית.
בכליה, שבה הדרישה האנרגטית גבוהה במיוחד והעומס המטבולי משתנה באופן תדיר, ציר NAD⁺–PARP–מיטוכונדריה🔋 מקבל משמעות קריטית. כאשר ציר זה יציב, התא מסוגל להתמודד עם שברים דו־גדיליים (DSB) כחלק ממערך תחזוקה רגיל. אך כאשר הוא קורס, כל אירוע נזק DNA הופך לאיום על עצם קיומו של התא.
פסקת המשך מצוינת למאמר, הממחישה את הקשר הישיר בין התזונה בצלחת לבין שבירת הגנום ותהליך ההצטלקות (פיברוזיס) של תאי הכליה דרך מטבוליט ה-TMAO:
🥩 מטבוליט ה-TMAO: הרעלן התזונתי ששובר את הגנום ישר מהצלחת שלך
במסגרת הנזקים שעלולים להתרחש בגרעין התא, לפגוע בחומר הגנטי ולשבור את שני גדיליו (DSB), בולט במיוחד מטבוליט הרסני בשם TMAO (Trimethylamine N-oxide). חומר זה אינו מיוצר בטעות, אלא מגיע אלינו כתוצר לוואי ישיר של התזונה היומיומית 🍳: כאשר אנו צורכים מזונות עשירים בכולין, לציטין וקרניטין – המצויים בעיקר בבשר אדום🍖, בשר בקר שמן🥩, איברים פנימיים🍔, ביצים🥚 ומוצרי חלב שמנים 🧀– חיידקי המעיים מפרקים אותם לתרכובת בשם TMA. זו נודדת לכבד, עוברת חמצון והופכת ל-TMAO, המשתחרר ישירות אל זרם הדם.
כאן ניצב אחד הפרדוקסים הטיפוליים הכי פחות מוכרים והכי מסוכנים ברפואת כליות (נפרולוגיה) כיום. הרופאים מקדישים לעבודתם הרבה זמן, לומדים, עובדים שעות ועוברים קורסים, אבל הם לא שולטים בדרך כלל במידע שמגיע אליהם, לפעמים דומה שמידע זה הוא הדהוד של כוחות כמו אלו שמהודהדים בערוץ 14 ובעיתון של מרים אדלסון כך: נפרולוגים רבים💉 מעודדים חולי כליה לצרוך חלבון ביצה (אלבומין) מתוך כוונה טובה – כדי למנוע איבוד שריר ותת-תזונה (Protein-Energy Wasting) מבלי להעמיס זרחן כמו בבשר. אולם, רצון טוב שמשולב באמונה טפלה 🙈, ומשענת שאינה קיימת במציאות🪽, מובילה גם נפרולוגים רבים 💉לאי ידיעה 🦜 וחוסר מודעות לכך שבביצים (ובמיוחד בחלמון) יש כמות עצומה של לציטין וכולין, שהם חומרי הגלם הישירים שחיידקי המעי הופכים ל-TMAO ההרסני לגנום.
🥚 מלכודת הביצים הכלייתית: המטבוליט ההרסני שמגיע דווקא מההמלצה הרפואית
תאכלו ביצים 🥚 זו דרישה שחושפת למעשה נושא חבוי שמסתתר תחת החלוק הלבן 🥼 פרדוקס רפואי דרמטי: נפרולוגים רבים מעודדים חולי כליה לצרוך חומרים, תזונה, ושאר דברים ששבים ונחשפים בעמודי העיתון הפנימיים כאסורים לנטילה, לפעמים מסרטנים, לפעמים התגלו כמקדמי מחלה אחרת, ובדרך כלל יצאו מהמחזור כח נמצא חומר סינטטי עם בשורה גדולה יותר. אולם, הציבור והממסד הרפואי לרוב אינם ערים לכך בכלל זה גם שביצים (ובפרט חלמון הביצה) הן מהמקורות העשירים והמרוכזים ביותר בטבע לכולין ולציטין – חומרי המקור המובהקים לייצור TMAO.
עבור תאי הכליה, רמות גבוהות של TMAO הן גזר דין מולקולרי חריף. כאשר ה-TMAO חודר לתאי האנדותל והנפרונים, הוא מחולל פקודה לייצור מוגבר וחסר בקרה של רדיקלים חופשיים (ROS) בתוך המיטוכונדריה. שטף ה-ROS הסוער הזה פולש אל גרעין התא ותוקף את קשרי הפוספט-סוכר השזורים בשני גדילי ה-DNA. התוצאה היא היווצרות ישירה ומתמשכת של שברים דו-גדיליים (DSB) קטסטרופליים, במיוחד באזורי הטלומרים הרגישים לחמצון.
מכיוון שהכליות הפגועות מתקשות לפנות את ה-TMAO מהגוף, אצל חולי כליות נוצר מעגל קסמים אכזרי: ההמלצה התזונתית לאכול יותר ביצים מקפיצה את רמות ה-TMAO בדם; הצטברות זו מייצרת עוד עקה חמצונית ועוד שברי DSB בגנום; הנזק הגנטי המתמשך מפעיל את אזעקת ה-p53 ודוחף את התאים הכלייתיים לסנסנציה (הזדקנות תאית); והפרשת הרעלים הדלקתיים (SASP) מאיצה את תהליך הפיברוזיס ואת קריסת הנפרונים שנותרו. כך, מה שנחשב המלצה רפואית, הנפרולוג אמר, הדיאטנים שלו הנחו, בשל בורות חסרת. אחריות כ"מזון בריא לכליות" בחדר הרופא💉, הופך למנוע מולקולרי ששובר את הגנום ומחריב את הרקמה הכלייתית מבפנים.

לצד המטבוליטים התזונתיים, הגנום הכלייתי חשוף גם לרעלנים סביבתיים כרוניים, ובראשם הדיאוקסינים (Dioxins). מזהמים תעשייתיים אלו נקשרים לקולטנים תאיים (AhR) ומחוללים סערה חמצונית מתמשכת בגרעין.
הדיאוקסין פוגע ישירות ביציבות האפיגנטית של הכרומטין ומייצר שטף בלתי פוסק של שברי DSB, המתישים את אנזימי התיקון ומאיצים את קריסת הנפרונים. כעזר מגיעים לרשותנו, כנזכר מתחת לצילום פמעלה, נבטי הברוקולי 🥦.
כך הקורא, המלומד, רואה בהדרגה, את מה שמערכת הבריאות הרגילה מפספסת!
התא בוחר בתיקון כאשר אנחנו בוחרים בתנאים המאפשרים אותו
מכאן נסגר המעגל הרעיוני של הפרק כולו: תיקון DNA אינו רק שאלה של חלבונים ותהליכים, אלא של סביבה ביולוגית שלמה — אפיגנטית, חמצונית ומטבולית — שמכריעה האם התא יבחר בתיקון והישרדות, או יגלוש לסנסנס, פיברוזיס וקריסה תפקודית.
מכאן נסגר המעגל הרעיוני של הפרק כולו, ושל סדרת מאמריי על נתיבי ההחלמה והפעלתם של מנגנוני ההגנה והשיקום של התא – רפואת העתיד.
תיקון ה־DNA אינו רק שאלה של חלבונים, אנזימים ומסלולים מולקולריים, אלא של הסביבה הביולוגית השלמה שבה חי התא. מצבו האפיגנטי, רמת העקה החמצונית, איכות התפקוד המיטוכונדריאלי, זמינות האנרגיה, העומס המטבולי והדלקתי, ואף תנאי החיים שאליהם נחשף האדם – כל אלה משפיעים על השאלה אם התא יצליח לגייס מנגנוני תיקון והתחדשות, או יידחף אל סנסנס, פיברוזיס ואובדן תפקוד.
הבנה זו מסמנת שינוי עמוק באופן שבו ראוי לחשוב על רפואה. במקרים רבים אין די בהתערבות תרופתית המכוונת למטרה ביולוגית אחת, משום ש
התא מקבל את החלטותיו מתוך רשת מורכבת של אותות ביולוגיים.
כאשר הסביבה התאית נותרת דלקתית, מחומצנת, דלה באנרגיה ועשירה בגורמי שחיקה, גם מנגנוני התיקון היעילים ביותר מתקשים למצות את יכולתם.
רפואת העתיד, ככל שהמחקר מתקדם, עשויה להתבסס יותר ויותר על יצירת תנאים המאפשרים לתא להפעיל את מנגנוני ההגנה, את צירי החלמה – והשיקום הטבעיים שלו. אין פירוש הדבר שתרופות יאבדו את מקומן; במחלות רבות הן מצילות חיים ומהוות מרכיב חיוני בטיפול. אולם לצד התרופות הולכת ומתבססת ההבנה ש
החלמה היא תהליך פעיל, הדורש יצירת סביבה ביולוגית המאפשרת לתא לבחור בתיקון ובהתחדשות.
במובן זה, החלמה איננה נס ואינה פעולה פסיבית. היא תוצאה של שילוב בין הידע המדעי לבין בחירות יומיומיות: תזונה מתאימה, פעילות גופנית, שינה מספקת, הפחתת גורמי שחיקה, איזון מטבולי, ושמירה על תנאים המאפשרים למנגנוני התיקון של הגוף לפעול במיטבם. ככל שנבין טוב יותר את מנגנוני ההחלמה הטבעיים של התא, כך נוכל לעבור מתפיסה המתמקדת בעיקר בטיפול במחלה, אל תפיסה השואפת ליצור את התנאים הביולוגיים המאפשרים בריאות והתחדשות.
צעדנו יחד בדיון על מנגנונים ביולוגיים אך בפועל התחלנו לראות מרחב, התבוננו וראינו כיצד הם מתחברים לסיפור אחד.
בתחילת הדרך אמנם היה לפנינו שבר דו־גדילי (DSB), שנראה כמו אירוע נקודתי. אבל ככל שמעמיקים, מתברר שהשבר הוא רק "קריאת השכמה". מיד אחריו מופיעים הכרומטין, הטלומרים, הסירטואינים (🍇🫐), המתילציה, המיטוכונדריה, ה־ROS, ה־NAD⁺, ובסופו של דבר אנו עומדים כמשה בהר נבו ומשתאים נוכח ההתכלות של הארץ המובטחת לא: "איך מתקנים שבר?", אלא באילו תנאים התא מסוגל בכלל לתקן אותו.
בכל תחנה בדרך ההחלמה מתגלה רובד עמוק יותר של אותה שאלה.
התחלנו כשאלה על תיקון DNA, ובמהלך הכתיבה הפך הקורא לצופה על התנאים המאפשרים תיקון DNA.
ככל שהעמקנו במנגנוני תיקון ה־DNA, התברר שהשאלה החשובה אינה כיצד התא מתקן את עצמו, אלא כיצד הוא שומר על היכולת לתקן. זהו מסע מן השבר אל התנאים המאפשרים התחדשות – מסע שמוביל מן המולקולה אל התא, מן התא אל הרקמה, ומן הרקמה אל אורח חייו של האדם.
חזרה לחלק א'
נשארו לך שאלות
🔬אשמח להשיב על כל שאלה
בבקשה לא להתקשר משום שזה פשוט לא מאפשר לי לעבוד – אנא השתמשו באמצעים שלפניכם
למען הסר ספק, חובת התייעצות עם רופא (המכיר לפרטים את מצבו הבריאותי הכללי של כל מטופל או שלך) לפני שימוש בכל תכשיר, מאכל, תמצית או ביצוע כל תרגיל. ירון מרגולין הוא רקדן ומבית המחול שלו בירושלים פרצה התורה כאשר נחשפה שיטת המחול שלו כבעלת יכולת מדהימה, באמצע שנות ה – 80 לרפא סרטן. המידע באתר של ירון מרגולין או באתר "לחיצות ההחלמה" (בפיסבוק או MARGOLINMETHOD.COM ), במאמר הנ"ל ובמאמרים של ירון מרגולין הם חומר למחשבה – פילוסופיה לא המלצה ולא הנחייה לציבור להשתמש או לחדול מלהשתמש בתרופות – אין במידע באתר זה או בכל אחד מהמאמרים תחליף להיוועצות עם מומחה מוכר המכיר לפרטים את מצבו הבריאותי הכללי שלך ושל משפחתך. מומלץ תמיד להתייעץ עם רופא מוסמך או רוקח בכל הנוגע בכאב, הרגשה רעה או למטרות ואופן השימוש, במזונות, משחות, תמציות ואפילו בתרגילים, או בתכשירים אחרים שנזכרים כאן.
physician (who knows in detail the general health of each patient or yours) before using any medicine, food, extract or any exercise. The information on Yaron Margolin's website or the "Healing Presses" website (on Facebook or YARONMARGOLIN.COM), in the above article and in Yaron Margolin's articles are material for thought – philosophy neither recommendation nor public guidance to use or cease to use drugs – no information on this site or anyone You should always consult with a qualified physician or pharmacist regarding pain, bad feeling, or goals and how to use foods, ointments, extracts and even exercises, or other remedies that are mentioned as such
מאמרים אחרונים
- שבר דו־גדילי ב־DNA: מסלולי התיקון, בחירת התא בין HR ל־NHEJ, והשלכות על סרטן, הזדקנות ומחלת הכליות / חלק ג' על ההחלמה ומה ניתן לעשות❓
- שבר דו־גדילי ב־DNA: מסלולי התיקון, בחירת התא בין HR ל־NHEJ, והשלכות על סרטן, הזדקנות ומחלת הכליות, חלק א' וחלק ב'/ ירון מרגולין
- צירי החלמה – מבוא למחשבה חדשה על בריאות והתחדשות מאת ירון מרגולין
- המסלולים הביולוגיים המשקמים את הגוף: פרספקטיבה חדשה על החלמה
- מנגנוני תיקון פנימיים בגוף: מודל אינטגרטיבי להבנת תפקוד הכליות והמערכת כולה
- 🇯🇵 腎機能と全身の健康に関わる生体調節メカニズムの概要
- 🔹 מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים🔹 את הגוף – חלק ב', על ההחלמה מאת ירון מרגולין
- המסלולים הביולוגיים שמשקמים את הגוף – חלק ג', על ההחלמה
- 🔷 המסלולים הביולוגיים שמשקמים את הגוף: p53 — שומר הגנום – חלק ד’ — ירון מרגולין
- חלק ה' – חלבוני ההחלמה: הרשת המולקולרית של הריפוי, ההתחדשות וההומאוסטזיס מאת ירון מרגולין
- חלק ו' על צירי ההחלמה ומה שמפעיל אותם – (signal architecture) מאת ירון מרגולין
- כבד שומני ❓- הכבד כמראה מערכתית: צירי מחלה, מול צירי החלמה – מבט אל רפואת העתיד
- 📘 The Biological Pathways That Restore the Body: A New Perspective on Recovery
- גמישות העורקים – רפואת העתיד – חלק א'
- עורקים גמישים חלק ב', הסבר מפורט על הצירים. הביולוגיה העמוקה של גמישות העורקים מאת ירון מרגולין
- קלוטו – האם קלוטו הוא מעיין הנעורים הזורם במערה מוסתרת וסודית?
- להחזיר את השליטה לכליה: תזונה נגד ספיגת היתר של הסוכר – איך לאזן את החלבון SGLT2
- תפקוד הכליה בשעות הלילה: השפעת המקצב הצירקדי על הפילטר הביולוגי
- טעם מר מעודד הפרשת רעלים מהכליות – רשימת צמחים בעלי טעם מר מאת ירון מרגולין
- הלשון של הכליות
- כיצד הכליה מטפלת בחלבונים
- אינסולין ותפקיד הכליות במטבוליזם ופינוי
- חולשת שרירים וירידה בתפקוד השרירים. כאבי שרירים, אבדן שריר, אבדן מסת שריר ומיופתיה.
- , יעד טיפולי חדש נגד מחלת הכליות
- ירידה בתפקוד הכליות – מה לעשות?
- בשר, כליות וזרימת דם מואצת: כיצד תזונה מהחי פוגעת במערכת הסינון של הגוף
- מה לאכול במצבי אי ספיקת כליות – מתכונים לדיאטה מאוזנת – טעימה להשתגע
- אי ספיקת כליות – מתכונים לדיאטה מאוזנת – טעימה להשתגע.
- זרימת דם כלייתית
- השפעות של הורמון הגדילה (GH) על תפקוד הכליות בבריאות ובמחלות כליות
- ריפוי פצעים וצמיחה של הכליה הנגדית לאחר כריתת כליה חד צדדית לצורך תרומה או טיפול
- לעשות?
- דיוקסינים במזון
- קדרה ואבדון
- Cauldron and Ruin
- אוכלוסיית המעי הגס חיידקים טובים, דיסביוזיס ו “שומר הגנום” גן p53
- מחסור בברזל
- מיומטי תנועה – התקווה החדשה לחולי כליות: תרכובות שמחקות את צירי ההחלמה של פעילות גופנית
- בשר, כליות וזרימת דם מואצת: כיצד תזונה מהחי פוגעת במערכת הסינון של הגוף
- תובנות חדשות אין להתחיל דיאליזה לפני בדיקת – מצבו של ציר FGF23-Klotho
- עיגול פינות מקשה על החלמת הכליות. מיומנו של מאסטר בהחלמת הכליות – הכליות לא סולחות על הזנחה:
- תובנות חדשות אי-ספיקת כליות תלויה במצבו של ציר FGF23-Klotho
- קלוטו – האם קלוטו הוא מעיין הנעורים הזורם במערה מוסתרת וסודית?
- תובנות חדשות אין להתחיל דיאליזה לפני בדיקת – מצבו של ציר FGF23-Klotho
- פחות מתים – טכניקת דיאליזה חדשה שבולמת את קצב המתים ומגבירה את ההישרדות בכשל כליות.
- שכיחות של דיכאון וחרדה בקרב חולי כליות
- המיקרוביום הכלייתי (האורוביום Urobiome) נחשף
- הערכות שונות במדידת אשלגן בפלזמה שוללות לפעמים שלא בצדק יתר אשלגן בה – היפרקלמיה פסאודו היפרקלמיה – Hyperkalemia
- אבנים בכליות – אוקסלטים, היפרוקסלוריה ויתר חומצה אוקסלית – רוצה להפעיל מנגנוני שיקום טבעיים בגוף האדם ללא תרופות?
- זרימת דם כלייתית
- השפעות של הורמון הגדילה (GH) על תפקוד הכליות בבריאות ובמחלות כליות
- סמן ביולוגי אמין (ביו-מרקר)
- למאמרי התכנית לשיקום הכליות – כאן
- ירידה בתפקוד הכליות – מה לעשות?
- ציר פה- כליות – פגיעה בבריאות הפה בקרב חולי כליות ו-CKD
- ⭐ התחדשות כלייתית: מקרה נדיר של ריפוי טבעי — ומה שהוא מלמד את כולנו
- סלניום Se התגלה כאנטי- אייג'ינג ומגן מפני מחלות כרוניות
- על ההחלמה – למה הגוף לא מחלים — וכיצד להחזיר לו את היכולת הטבעית לעשות זאת
- קרקר מדהים מכוסמת ירוקה – מתכון
- על ההחלמה – למה הגוף לא מחלים — וכיצד להחזיר לו את היכולת הטבעית לעשות זאת
- שינה חשובה להחלמה – 🌜זמן הריפוי החשוב מתרחש בלילה
- ניקוי כבד טבעי – הדרך הפשוטה לבריאות מתחדשת
- תורת הנפש לאפלטון – חלק1
- תורת הנפש של אפלטון – גיבור הרוח האנושית חלק2.
- תורת הנפש של אפלטון – הידיעה חלק3
- שפינוזה כממשיך דרכו של אפלטון – תורת הנפש חלק4
- Naturalne uzdrowienie: droga powrotu do zdrowia z ciężkiej choroby przewlekłej — niewydolności nerekUzdrowienie jest możliwe. Nadzieja jest prawdziwa.
- Шлях до одужання від тяжкого хронічного захворюванняВісім основних принципів Ярона Марголіна, майстра з відновлення
- principes fondamentaux de la guérison naturelle de Yaron Margolin
- Естественное исцеление: путь к выздоровлению от тяжелого хронического заболевания Восемь основных принципов Ярона Марголина, мастера по восстановлению функции почек
- uzdrowienie: droga powrotu do zdrowia z ciężkiej choroby przewlekłej — niewydolności nerekUzdrowienie jest możliwe. Nadzieja jest prawdziwa.
- Шлях до одужання від тяжкого хронічного захворюванняВісім основних принципів Ярона Марголіна, майстра з відновлення функції нирок
- Doğal Şifa: Zorlayıcı Kronik Hastalıklardan İyileşmenin Yolu – Yaron Margolin, Böbrek İyileşmesi Ustası’nın Sekiz Temel İlkesi
- หลักการฟื้นฟูไตทั้งแปดประการโดย ยารอน มาร์โกลิน –
- Natürliche Heilung – Der Weg zur Heilung von chronischen ErkrankungenDie acht Grundprinzipien von Yaron Margolin, Meister der Nierenheilung
- Learn the 8 Principles That Revive Kidney Function
- Sanación natural: El camino hacia la recuperación de una enfermedad crónica grave – Ocho principios fundamentales de Yaron Margolin, maestro en la recuperación renal
- Vous pouvez guérir de l’insuffisance rénale – Les huit principes fondamentaux de la guérison naturelle de Yaron Margolin
- Weg zur Heilung von chronischen ErkrankungenDie acht Grundprinzipien von Yaron Margolin, Meister der Nierenheilung
- Learn the 8 Principles That Revive Kidney Function
- Sanación natural: El camino hacia la recuperación de una enfermedad crónica grave – Ocho principios fundamentales de Yaron Margolin, maestro en la recuperación renal
- Vous pouvez guérir de l’insuffisance rénale – Les huit principes fondamentaux de la guérison naturelle de Yaron Margolin
- מסלולי ההחלמה -המסלולים הביולוגיים שמשקמים את הגוף – חלק א' מאת ירון מרגולין.
- בשר, כליות וזרימת דם מואצת: כיצד תזונה מהחי פוגעת במערכת הסינון של הגוף
- באיזה אופן תהליך המתילציה משפיע על ההחלמה מחלות כרוניות
- איך לא להגיע לדיאליזה
- "הצלבת איברים – הדרך להבנתה של המורכבות האדירה של מחלת כליות קשה מאוד" כאשר מסלולי ההחלמה מושבתים – כאן..
- פחות מתים – טכניקת דיאליזה חדשה שבולמת את קצב המתים ומגבירה את ההישרדות בכשל כליות.
- ציר פה- כליות – פגיעה בבריאות הפה בקרב חולי כליות ו-CKD
- רככת היפופוספטמית (זרחן נמוך מהנורמה) – מחלת כליות כרונית – או הפרעת עצם מינרלית
- מלטונין הוכח שהורמון השינה נוגד חמצון רב עוצמה ויעיל כתרופה בקרב חולי כליה
- 8 חסרים תזונתיים שכיחים
- מזון כתרופה
- עוצמתם של נבטי הברוקולי כתרופה לחולי כליה
- התערבויות בניהול רגשות, מחשבות ואורח חיים כדי להתמודד עם מגיפה של מחלות כליתיות
- תפקוד הכליה כבלוטה הורמונלית: סקירה מדעית
- אינסולין ותפקיד הכליות במטבוליזם ופינוי
- הפשטות שמרפאה: איך תזונה חיה מחזירה גמישות לעורקים
- ניקוי כבד טבעי – הדרך הפשוטה לבריאות מתחדשת
- גמישות העורקים
- שלושת התוספים החשובים ביותר לאדם מעל גיל 55 — גישה טבעית וזהירה
- קדרה ואבדון
- התערבויות בניהול רגשות, מחשבות ואורח חיים כדי להתמודד עם מגיפה של מחלות כליתיות
- פורינים חוץ תאיים ותחזוקת הכליות
- מחלת כליות – נתנת לשינוי – לשם כך יש לזהות אותה מוקדם ככל האפשר – הקסטסרופה!
- להפעיל תהליך התאוששות בסוכרת ואי ספיקת כליות
- הקשר של כבד שומני, ומחלת כליות כרונית מחקר חדש
- אנמיה – מחסור בברזל, כשל טיפול והיבטים ביולוגיים חשובים של מחסור בברזל ודרכים חדשות של הפעלת צירי ההחלמה בגוף האדם – מחקר חדש.
- למה חסר לי ברזל – כאן
- לא עומד לי דיקום הטסטוסטרון והאון – בלי תרופות, עם דרך אמיתית
- צרבת כרונית – רוצה להפעיל תהליך התאוששות ללא תרופות?
- ירידה בתפקוד הכליות – מה לעשות?
- חומצה אלפא לינולנית משפרת את תוחלת החיים – צריכה גבוהה של חומצות שומן מצמחים נחשפה
- ריפוי טבעי: הדרך לאפשרות ריפוי ממחלה כרונית קשה שמונה עקרונות היסוד של ירון מרגולין
- חילוף חומרים אנרגטי, איזון רקמת השומן ובקרת תיאבון
- ירקות ירוקי עלים לרוב טובים לבריאותנו, לפעמים הם לא – רוצה לדעת מתי כדאי לצרוך עלים ירוקים?
רפואת העתיד – מחקרים מתקדמים כיום מצביעים שוב ושוב על דרך פשוטה "להדליק" באופן ממוקד את חלבוני ההחלמה וצירי השיקום הטבעיים שבגוף האדם. אחד הנושאים המרתקים ביותר כיום ממוקד ב HR NHEJ.זהו אחד השדות הלוהטים ביותר כיום בביולוגיה מולקולרית וברפואה, שמבקשת שינוי. לא עוד תחזוקה וחיי אומללות: החלמה.מכיוון שה HR NHEJ מהווה את הבסיס להצלחה או לכישלון של עריכה גנטית (כמו CRISPR) ולפיתוח דרכי החלמה פורצות דרך.ה HR NHEJ מוביל ישירות אל נושא צירי ההחלמה, חלבוני החלמה, חסימת מסלולי מחלה, והפחתת סטרס חימצוני – צירים אלו הם בעצם נתיבי החלמה לא־תרופתיים שעשויים להפחית את עומס הנזק ל־DNA ולתמוך בסביבה התאית הנחוצה לתיקון תקין: הפחתת עקה חמצונית, שיפור תפקוד מיטוכונדריאלי, שימור מאגרי NAD⁺, תמיכה בפעילות סירטואינים כגון SIRT1 ו־SIRT6, שמירה על יציבות אפיגנטית, והגנה על טלומרים. במילים אחרות, במקום "להכריח" את התא לתקן, אפשר לנסות לשפר את התנאים שבתוכם מנגנוני התיקון פועלים. מאמר בעל שבר דו-גדילי (DSB) בסדרת צירי ההחלמה מאת ירון מרגולין